All published articles of this journal are available on ScienceDirect.

Listeriolysin O, but not Murine E-cadherin, is Involved in Invasion of Listeria monocytogenes into Murine Liver Parenchymal Cells
Abstract
Human E-cadherin and listeriolysin O (LLO) are involved in invasion of Listeria monocytogenes into human liver parenchymal cells (LPC). Yet, it remains to be determined whether murine E-cadherin and LLO participate in invasion of L. monocytogenes into murine LPC. In the present study, involvement of murine E-cadherin and LLO in invasion of L. monocytogenes into murine LPC was investigated. Murine E-cadherin was expressed on murine LPC, but the expression became undetectable by insertion of transgene of Simian virus 40 large T antigen. Although invasion of L. monocytogenes into murine LPC was found regardless of murine E-cadherin expression, infection rate of L. monocytogenes being unable to secrete LLO was lower than that of L. monocytogenes being capable of secreting LLO. Our RESULTS verify that invasion of L. monocytogenes into murine LPC occurs independently of murine E-cadherin and indicate that LLO participates in invasion of L. monocytogenes into murine LPC.
Listeria monocytogenes is a facultative intracellular bacterium which can survive and replicate in professional phagocytes such as macrophages (Mφ) [1]. Listeriolysin O (LLO) encoded by hly gene is one of the most important virulence factors of L. monocytogenes [2]. LLO allows L. monocytogenes to escape from the phagosome into the cytosol in Mφ [3-5]. A vast majority of L. monocytogenes are trapped in the liver immediately after systemic infection [6] and liver parenchymal cells (LPC) serve as a habitat of this bacterium [7], suggesting that L. monocytogenes invades LPC using any entry factor(s). Although LLO has been shown to be involved in invasion of L. monocytogenes into human LPC [8, 9], it remains to be determined whether LLO participates in invasion of this bacterium into murine LPC.
E-cadherin is an intercellular adhesion molecule highly expressed on basolateral membrane of intestinal epithelial cells (IEC) as well as LPC [10-13]. Because (i) L. monocytogenes expresses internalin A (InlA) encoded by inlA gene [10, 14], (ii) InlA is one of the ligands for human E-cadherin [10, 15], (iii) L. monocytogenes lacking inlA gene is unable to invade human IEC [16, 17] and (iv) L. monocytogenes passes through the intestinal barrier of transgenic mice expressing human E-cadherin in InlA-dependent manner although no invasion of L. monocytogenes is found in normal mice [18], interaction between InlA and E-cadherin plays a pivotal role in invasion of L. monocytogenes into human IEC [10, 14-17].
Similar to human, E-cadherin is expressed on murine LPC as well as IEC [12, 13]. However, because an amino acid sequence of murine E-cadherin considerably differs from that of human E-cadherin, murine E-cadherin is unable to bind to InlA [19]. Yet, L. monocytogenes is isolated not only from the mesenteric lymph nodes, but also from the liver and the spleen after p.o. infection in normal mice [20-22]. It is therefore possible that unknown mechanism exists in invasion of L. monocytogenes into murine LPC and IEC. Indeed, ligand(s) other than InlA which can interact with E-cadherin has been identified in both human and mice [23-25]. Therefore, we raise the question of whether ligand(s) for murine E-cadherin other than InlA participate in invasion of L. monocytogenes.
In the present study, we reevaluated whether murine E-cadherin is not indeed involved in invasion of L. monocytogenes into murine LPC using LPC expressing and lacking murine E-cadherin, and examined whether LLO is involved in invasion of this bacterium into murine LPC.
We first compared the surface expression of murine E-cadherin on AII (kindly provided by Dr. Peter Lösser (Robert Koch Institute)) derived from p53 knockout mice [26] and HepSV40 (kindly provided by Dr. Peter Lösser (Robert Koch Institute)) derived from Simian virus (SV) 40 large T antigen transgenic mice [27] by flow cytometry. AII and HepSV40 were stained with carboxyfluorescein-conjugated anti-mouse E-cadherin mAb (Clone: 114420; R&D Systems, Minneapolis, MN). After washing with PBS containing 0.1 % bovine serum albumin (Wako Pure Chemical Industries, Osaka, Japan) and 0.1 % sodium azide (Wako Pure Chemical Industries), cells were acquired by FACSCalibur® (BD Biosciences, Mountain View, CA) and murine E-cadherin surface expression was analyzed with Flow Jo software (version 7.6.5; Tomy Digital Biology, Tokyo, Japan). As we expected, the majority of AII expressed murine E-cadherin (Fig. (1)). Surprisingly, murine E-cadherin-expressing cells were only marginal in HepSV40.
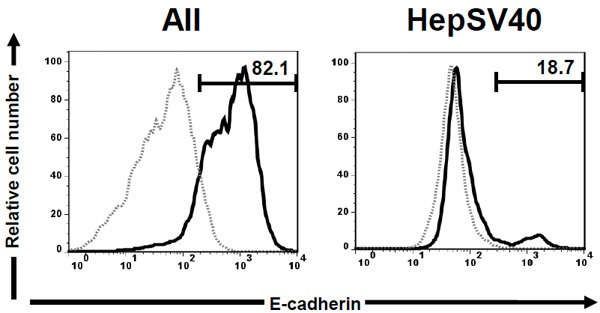
Cell surface expression of murine E-cadherin on AII and HepSV40. AII and HepSV40 were stained with carboxyfluorescein- conjugated anti-mouse E-cadherin mAb and the E-cadherin surface expression was analyzed by flow cytometry. The profiles of E-cadherin are displayed as histograms. Dotted and solid lines represent LPC lines unstained and stained with anti-mouse E-cadherin mAb. Numbers in histograms represent percentages of E-cadherin+ cells. Representative staining patterns from 2 independent experiments are shown.
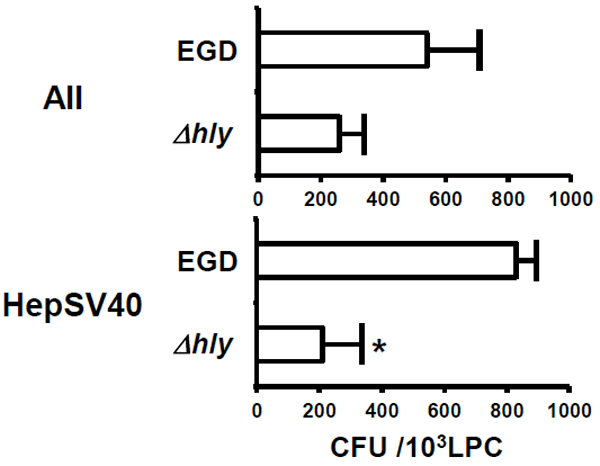
CFU in AII and HepSV40 after L. monocytogenes infection. AII and HepSV40 were incubated with L. monocytogenes infection (strains EGD or Δhly) in RPMI 1640 suplemented with 10 % fetal calf serum (complete medium; CM) for 1 h. After removing their culture supernatants, the LPC lines were washed 3 times with CM containing 10 µg/ml of gentamicin to kill extracellular L. monocytogenes infection. Immediately after washing with CM, a portion of cells were plated on tryptic soy agar plate after sonication, and the CFU in LPC were enumerated by plate counts after 48 h incubation. Representative data from 3 independent experiments are shown. The statistical significance was determined by Student t-test and p value of < 0.05 was regarded as significant. *, p < 0.05 : EGD vsΔhly.
To examine whether murine E-cadherin is not indeed involved in invasion of L. monocytogenes into murine LPC, AII (5 × 104 cells) expressing murine E-cadherin and HepSV40 (2 × 105 cells) lacking murine E-cadherin were incubated with 5 × 105 colony-forming units (CFU) and 2 × 106 CFU of L. monocytogenes (strains EGD and Δhly), respectively for 1 h and the CFU in these cells were determined as described previously [28]. Considerable numbers of L. monocytogenes were detected in both AII and HepSV40, and the numbers were comparable (Fig. (2)). The number of strain EGD in HepSV40 was significantly higher than that of strain Δhly. Similarly, the number of strain EGD in AII was considerably, though not significantly, higher than that of strain Δhly (Fig. (2)).
We show here the involvement of murine E-cadherin and LLO in invasion of L. monocytogenes into murine LPC. The number of strain EGD in HepSV40 was slightly, though not significantly, higher than that in AII, despite the fact that the number of E-cadherin+ cells was markedly higher in AII as compared to HepSV40. These RESULTS strongly indicate that murine E-cadherin is not involved in invasion of L. monocytogenes into murine LPC. Because (i) internalin B (InlB) is one of the ligands for hepatocyte growth factor receptor (HGF-R / Met) [14, 16], (ii) Met and InlB are highly expressed on murine LPC and L. monocytogenes, respectively [14, 29] and (iii) the interaction between InlB and Met has been shown to play a central role in invasion of L. monocytogenes into human and murine IEC [30-34], we consider it likely that L. monocytogenes invades murine LPC through interaction between Met and InlB.
Numbers of strain Δhly in AII and HepSV40 were markedly lower than those of strain EGD. Because strain Δhly is unable to secrete LLO [35] and because LLO has been shown to participate in invasion of L. monocytogenes into human LPC [8, 9], we assume that LLO is involved in invasion of L. monocytogenes into murine LPC.
Although E-cadherin is normally expressed on the surface of LPC [11, 12], HepSV40 lacked surface expression of E-cadherin. Consistent with this, Rip1Tag2 mice which are SV40 large T-antigen transgenic mice also lack surface expression of E-cadherin [36]. We therefore consider it likely that SV40 down-regulates E-cadherin surface expression on LPC.
In conclusion, we verified that murine E-cadherin was not involved in invasion of L. monocytogenes into murine LPC and obtained the first evidence that LLO participates, at least in part, in invasion of L. monocytogenes into murine LPC. Although we were unable to determine the molecule(s) which participate in invasion of L. monocytogenes into murine LPC, we consider it likely that interaction between InlB and Met is involved in this mechanism. As shown here, characteristic features (e.g. E-cadherin surface expression) of cells are dramatically changed by insertion of transgene. It is therefore possible that the insertion of transgene influences on the data, by which the conclusion could be changed. Therefore, the data obtained from experiment using transgenic mice should be reevaluated and the conclusion should be drawn with care.
ABBREVIATIONS
CONFLICT OF INTEREST
The authors confirm that this article content has no conflict of interest.
ACKNOWLEDGEMENTS
This work was supported by a Grant-in-Aid for Scientific Research (22300261 to M.E. and 22590388 to Y.E.) from the Japan Society for the Promotion of Science.

