All published articles of this journal are available on ScienceDirect.

Bioinformatic Studies, Experimental Validation of Phytase Production and Optimization of Fermentation Conditions for Enhancing Phytase Enzyme Production by Different Microorganisms under Solid-State Fermentation
Abstract
Background:
Phytase is an essential enzyme necessary for the digestive process. It is a natural enzyme found in plant materials. It prevents bad effect of phytic acid on protein and energy utilization. Phytase frees the bound minerals such as phosphorus, calcium, zinc, iron, magnesium and manganese from the phytic acid molecule providing essential minerals available for healthy nutrition. This study depends on converting food processing waste into highly valuable products. Optimizing the fermentation conditions for enhancing high phytase production with low cost was the objective of this research.
Methods:
A bibliographical survey was carried out to select the most fungul producers of phytase from fungal species deposited in NCBI database. Phytases of the selected organisms were analyzed in the UNIPROT database and their protein sequences were submitted to multiple sequence alignments using Clustal Omega and visualized using Jalview program. Experimental studies using five fungal strains of Aspergillus.ssp on wheat bran under Solid-State Fermentation carried out. Comparisons were made for phytase production. A. awamori NRC- F18 as the best phytase producer-strain cultured on different types of treated wastes followed by optimizing the fermentation conditions for enhancing phytase production using rice straw as the best substrate, which provides the highest phytase production. Thermostability of crude enzyme was studied. Statistical analyses were performed using SPSS at P < 0.05 or P < 0.01.
Results:
Bioinformatic studies predicted the most producer species and explained the difference in activity of phytases produced from different species, although they have the same function. All phytases of the selected fungal species from the database NCBI have highly conserved amino acid sequences; there are 88 identical positions; 135 similar positions, but the identity percentage was 16.858%. Experimental studies using five fungal strains of Aspergillus ssp. on wheat bran revealed optimum conditions for phytase production by A. awamori NRC- F18, which cultured on different types of treated wastes. A considerably higher phytase production was obtained using rice straw as substrate 424.66± 2.92 IU /g at pH 6 (371.883± 0.822 IU /g), after 144 hrs of incubation at 30°C. The maximum enzyme activity observed when solid: moisture was 1:4; Inoculum concentration 2mg/5g (418.363± 16.709 IU /g) and substrate concentration 4.5% (277.39± 12.05 IU /g). Glucose and Ammonium acetate were the best carbon and nitrogen sources that enhanced phytase production from A. awamori NRC- F18. The obtained phytase was found to be thermostable and the maximum temperature at which phytase still active was 80°C.
Conclusion:
Bioinformatic studies predicted the most producer species. Experimental study revealed that A.awamori NRC- F18 was the best Phytase -producer strain. Solid state fermentation was a good method; pretreatment of agriculture residues as rice straw was useful for less expensive phytase production, which was thermostable. A. awamori NRC- F18 can be used in the industrial production of phytase.
1. INTRODUCTION
Phytases have a potential role in the food industries as it reduces the phytic acid, which binds cations (Ca2+, Fe2+, Zn2+, Mg2+, and P2+) forming insoluble complexes in the intestinal tract causing mineral malabsorption content [1, 2]. Phytases are classified structurally into four groups; the histidine acid phosphatase (HAP) superfamily, β-propeller phytase (βPP), purple acid phosphatase (metalloenzyme-PAP), and protein tyrosine phosphatase (cysteine-phytase) [3]. All groups have catalytic mechanisms. The first group of phytases (HAP superfamily) is the most studied group [4]. HAP group catalyzed hydrolysis of substrate through nucleophilic attack of catalytic H on phosphomonoester and the hydrolysis of a covalent phosphohistidine intermediate with releasing of H [5, 6]. The HAP phytases possess the active site motif related to the catalysis and HD for substrate binding. Phytase has a vital role in human nutrition to overcome the negative effects of phytic acid found in most cereals and legumes which are rich in protein and fat but they have antinutritional factors like phytic acid which forms complexes with proteins, resulting in a decrease of their solubility and digestibility, leading to reduce their nutritive value. In addition, phytic acid chelates vitamin B3 causing Pellagra, a vitamin deficiency disease. Phytase is an important supplement in poultry rations which involved components contain phytaic acid as conola seeds and wheat bran to overcome the problems of phytate which acts as an antinutritional agent in monogastric animals by chelating important minerals in the ration as well as protein and vitamins [4]. Therefore, the reduction of the phytate content in seed meal via its enzymatic addition is demand [7, 8]. Fungal isolates belonging to the genera Mucor, Penicillium, Rhizopus and Aspergillus have been identified as active producers of phytases [9, 10]. The most remarkable one and a commercial source for phytase is Aspergillus niger [11, 12]. The enzymatic degradation of phytic acid will not produce toxic by-products; hence, it is considered environmentally friendly [13]. In view of the increasing demand for phytase, cost must be taken into consideration when applying microorganisms for commercial phytase production. Using inexpensive carbon, high producing fungal strains and suitable fermentation systems are factors for low-cost phytase production. The production of phytase from fungi has been achieved using three different cultivation methods, namely, solid-state, semisolid and submerged fermentation [14, 15]. Solid-State Fermentation (SSF) system provides a great deal of interest in the present years because it offers several economical and practical advantages, including high product concentration, improved product recovery, simple cultivation equipment, and lower plant operational cost [16-18]. Low cost phytase depends on using an inexpensive carbon source. Research for efficient nonexpensive carbon sources must be taken into consideration. This work aimed to select some fungi which have the potential for phytase production through in silico studies of the physicochemical properties and the secretion parameters of the predicted phytases. Finally, the experimental validation by studying the ability of different microorganisms to produce phytase enzyme using Solid -State fermentation (SSF), choosing the best phytase producing-microorganism and studying the effect of media ingredients on phytase enzyme production. Also, this report aims to produce low-cost phytase enzyme by using inexpensive media, high producing fungal strains, a suitable fermentation system and optimal conditions for phytase production during SSF on food processing waste.
2. MATERIALS AND METHODS
2.1. Bioinformatic Analysis
A bibliographical survey was carried out to select the most fungul producers of phytase. After the initial study, six species were selected from fungal species which deposited at the database National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/). For this work, phytases were analyzed in the UNIPROT database (UniProtKB/Swiss-Prot). Protein sequences of phytases for the selected organisms were obtained in FASTA format with accession numbers:(3-phytase A: P34752.1 [A. niger]; 3-phytase B: P34754.1[A.niger]; 3-phytaseA:P34753.1 [A.awa mori];3-phytaseB:P34755.1[A.awamori]; 3-phytase A: O00092.1 [A. fumigatus Af293]; 3-phytase A: Q9C1T1.1[A. oryzae RIB40]) using UniProtKB/Swiss-Prot [19].
2.2. Multiple Sequence Alignments
The main idea behind building a multiple sequence alignment is to put similar amino acids in the same column if they contain the same criterion. The sequences of the phytases under study (in FASTA format) were submitted to multiple sequence alignments using Clustal Omega (https://www.ebi .ac.uk/Tools/msa/clustalo/) to identify conserved sites among the selected phytases. Then the multiple sequence alignments were visualized using the Jalview program Version: 2.11.1.3www.jalview.org/faq for analysis of residue conservation. Bioinformatics tools followed by experimental validation of phytase production by five fungal strains of Aspergillus ssp, which compared to T. viride as well as S. cerevisiae.
2.3. Experimental Validation of Phytase Production
2.3.1. Agro-industrial Substrates
Wheat bran, sugar beet pulp, cotton seeds, sorghum flour and cornflour were purchased from the local market. Orange and pomegranate peels as a food industry wastes, rice straw from local filed, El Shariqia Governorate, and sugar bagasse pith from Dshna sugar factory, Egyptian of sugar and integrated industries company were purchased.
2.3.2. Pretreatment of Rice Straw and Sugar Cane Bagasse Pith
Crushed sugar cane bagasse pith and rice straw were pre-treated with 1.5% sodium hydroxide at solid: liquid ratio 1: 20 at 121oC for 30 minutes. After cooling, the alkali was neutralized with HCl to pH 6. The solid treated substrates were filtered through cheeseclothes washed several times with water and finally drying in the oven at 70oC till constant weight was achieved.
2.3.3. Microorganisms and Inoculum Preparation
Non-mycotoxin producing fungal strains, namely Aspergillus oryzae F -923; A. oryzae F -937; A. niger F-258; A.awamori NRC- F18.; A. fumigates NRCF-113 and T. viride NRCF-107 as well as S. cerevisiae F-307 were obtained from Microbial Chemistry Lab. National Research Center, Dokki, Cairo, Egypt and maintained on potato dextrose agar (PDA). Pure cultures stored at 4oC and sub-cultured monthly. Inoculum was made from 5 days old PDA cultures. The inoculum (containing 106-107 spores) was suspended in YPM broth. The suspension, when necessary, was diluted with sterile water to give a spore count within the predetermined range. One ml of inoculums was added to each cooled sterilized (autoclaving at 121oC for 15 minutes) media.
2.3.4. Evaluation of Different Types of Microorganisms for Phytase Production
Fungal strains screening for phytase production was studied in 250 ml Erlenmeyer flasks containing 5g of wheat bran moistened with distilled water at solid liquid ratio 1:2 autoclaved at 121oC for 20 min and 15 psi. One milliliter of spore suspension from each fungal was used as inoculum for each flask. The cultures were incubated statically at 30oC for 5 days. All experiments were carried out in a triplicate manner [11, 12, 20-22].
2.3.5. Evaluation of Different Types of Wastes for Phytase Production
The selected fungal A.awamori NRC- F18 strain was cultured on nine substrates, namely sugar beet pulp, sorghum flour, cornflour, orange and pomegranate peels, wheat bran and defatted cotton seeds moistened with distilled water to moisture levels 1:2 (w/v) except rice straw and sugar cane bagasse pith moistened with the solution containing 4 g di-ammonium hydrogen phosphate and 0.5 g magnesium sulfate /L. The inoculated cooled sterilized flasks were incubated for 5 days at 30oC [23-26].
2.3.6. Effect of Nitrogen Source
Ten different nitrogen sources, namely urea, ammonium hydrogen phosphate, diammonium hydrogen citrate, ammonium oxalate, potassium nitrate, sodium nitrate, ammonium nitrate, ammonium acetate and ammonium sulphate and ammonium phosphate at equivalent nitrogen level were tested for selection of the more suitable nitrogen source for enzyme production [1, 27, 28].
2.3.7. Effect of Carbon Source
The influence of addition soluble carbon sources on phytase production in the SSF was studied. Rice straw medium was supplemented with different twelve carbon sources, namely maltose, sorbose, cellobiose, dextran, galactose, arabinose, glucose, xylose, lactose, mannitol, and fructose at a level of 1:5 w/w. The medium without a carbon source served as control [29].
2.3.8. Effect of Initial pH on Phytase Production
The effect of medium pH on phytase production was studied by adjusting the initial pH of fermentation medium to different values from 3 to 7.5 with0.1 M acetate buffer [6].
2.3.9. Effect of Substrate % to Fermentation Container Volume on Phytase Production
Rice straw medium was prepared with different concentrations (1.5% to 5.5% w/v) to study the influence of substrate % to fermentation container on phytase production in the SSF [30].
2.3.10. Effect of Moisture Level on Phytase Production
Different moisture levels ranged 1:1 - 1:5 w/v solid-liquid ratio were used to study the influence of moisture level on phytase production in the SSF [31, 32].
2.3.11. Effect of Inoculums Size on Phytase Production
Different Inoculum sizes ranged (0.5-2.5ml/5g substrate were used to study the influence of inoculums size on phytase production under SSF [1].
2.3.12. Effect of Fermentation Period on Phytase Production
The effect of fermentation time on phytase production was studied by incubating fungus in a solid state cultivation medium for 7 days and periodically testing the enzyme activity [1, 33].
2.3.13. Extraction of Enzymes
After the incubation period, the extraction solvent was added to each flask of the fermented cultures in ratio (10:1) (v/w) solvent to the substrate and 0.1%Tween 80. The flasks were shaken at 150 rpm for 30 min at 30oC on a rotary shaker. Then the content of each flask was filtered through Whatman filter paper No.3 to obtain a clear filtrate. The collected filtrate was used as a crude enzyme extract for assaying the activity of phytase.
2.3.14. Phytase Assay
Phytase activity was assayed by measuring the amount of inorganic phosphorus released from sodium phytate solution. Stock solutions: 10 mM ammonium molybdate, 1 M citric acid and 2.5 M sulfuric acid were prepared. Reaction mixture prepared by adding 0.5 ml (filtrate) diluted extract to 0.5ml sodium phytate 0.1% (made up in 200 mM acetate buffer pH 5.5) incubated at 50oC for 15 minutes. 4ml of freshly prepared constituents of above prepared 10 mM ammonium molybdate, 2.5 M sulfuric acid and acetone at ratio 1: 1: 2, respectively, followed by vortex before addition of 0.4 ml citric acid. The absorbance was measured at 420 nm. One unit of enzyme activity was defined as the amount of phytase required to release one micromole of inorganic phosphorus per minute under the assay conditions. Standard curve was performed using di-potassium hydrogen phosphate as a source of phosphorus with working concentration ranging from 10 to 100μM [27, 34].
2.4. Statistical Analysis
Statistical analyses were performed using the Statistical Package for Social Sciences (SPSS) software and Arab processor in social statistics. Descriptive statistics such as means and Standard Deviations (SD) were calculated for nonparametric variables were tested by Chi-square tests. Differences were considered statistically significant at P < 0.05 or P < 0.01 [38, 39].
3. RESULTS
3.1. In Silico Analysis
There were 33.773 phytases in UNIPROT Unreviewed (Tr-EMBL) data base and 23 phytases in UNIPROT reviewed (Swiss-Prot). 23 phytases in total were reported as revised or cured. Six phytases of cured phytases were selected and the distribution was as follows: two for A. awamori; two for A. niger and one for each species A. oryzae and A.fumigatus. The multiple alignment (using CLUSTAL_O) between the amino acid sequences of the phytases were reported for Aspergillus spp enabled the identification of a highly conserved sequence (Fig. 1).

(*) A star indicates an entirely conserved column; (:) A colon indicates columns where all the residues have roughly the same size and the same hydropathy; (.) A period indicates columns where the size OR the hydropathy has been preserved in the course of evolution.
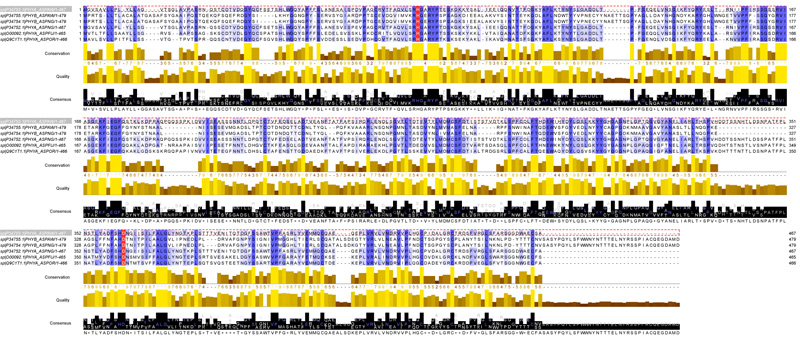
Blue columns: conserved amino acids in phytase sequences with 100% identity. Red columns: the position of active site in phytase sequences. Height of the letters in the Web Logo: indicates the degree of conservation at specific locations.
The result presented in Fig. (2) revealed that the MSA program used in UniportKB is Clustal.O; there are 88 identical positions and the identity percentage is 16.858%. There are 135 similar positions between these sequences. The first conserved aromatic amino acid in all selected organisms is Y(tyrosine). The first conserved amino acids G (glycine) and P (proline) in all selected organisms are at positions 43 and 59, respectively. There are 13 conserved columns of amino acid G and 8 conserved columns of amino acid P. The first conserved amino acids C (cysteines) in all selected organisms is C77. There are 5 conserved columns of amino acid C. The first conserved amino acids H(His tidine), S(serine) in all selected organisms are H88 and S30. There are 2 conserved columns of amino acid H and 7 conserved columns of amino acid S. The first conserved amino acids K (Lysine), R (Arginine), D (Aspartic Acid), E (Glutamic Acid) in all selected organisms are K189; R87; D242 and E192. There are 1 conserved column of amino acid K; 6 conserved columns of amino acid R; 2 conserved columns of amino acid D and 4 conserved columns of amino acid E. The first conserved amino acids L (Leucines) in all selected organisms is L7. There are 11 conserved columns of amino acid L. The first conserved amino acids W (tryptophans), F (phenylalanine), Y (tyrosine) in all selected organisms are W307; F122 and Y44. There are 1 conserved column of amino acid W; 6 conserved columns of amino acid F and 7 conserved columns of amino acid Y.
Table 1 revealed that phytases of all selected species have sequence length between (465-479). 3-phytase A [A. niger]: P34752.1 and 3-phytase A [A. awamori]: P34753.1 have the same Sequence Length 467. The active site at positions H82 and D362 in two sequences. 3-phytase B [A.niger]: P34754.1 and 3-phytase B [A.awamori]: P34755.1 have the same sequence length 479. The active site at the positions H81 and D338 in two sequences. Sequence length of 3-phytase A [A.fumigatus]: O00092.1 was 465 and the active site was at the position H81 and D360 while Sequence Length of 3-phytase A [A. oryzae]: Q9C1T1.1. was 466 and the active site was at the positions H81 and D361.
| Organisms | Protein names | Accession numbers | Sequence Length | Mass (Da) | Active site |
|---|---|---|---|---|---|
| A. fumigatus | 3-phytase A | O00092.1 | 465 | 50.836 | H81/D360 |
| A.niger | 3-phytase A | P34752.1 | 467 | 51.086 | H82/D362 |
| A. awamori | 3-phytase B | P34755.1 | 479 | 52.678 | H82/D338 |
| A.awamori | 3-phytase A | P34753.1 | 467 | 51.075 | H82/D362 |
| A. oryzae | 3-phytase A | Q9C1T1.1 | 466 | 51.257 | H81/ D361 |
| A. niger | 3-phytase B | P34754 | 479 | 52.612 | H82/D338 |
3.2. Experimental Validation of Phytase Production
3.2.1. Evaluation of Different Fungal Strains for Phytase Production on Wheat Bran Under Solid State Fermentation
Screening of different fungal strains for phytase production was carried out on 5g of wheat bran moistened with distilled water at solid liquid ratio 1:2 for each microorganism.
The results were statistically analyzed, presented in Table 2 and showed that there was a significant difference between treatments and the tested fungi A. awamori NRC- F18 provide the highest phytase activity (143.67±7.30IU/g) followed by A. niger F-258 (110.62± 9.85IU/g) then A. oryzae F -923(66.56 ± 1.70IU/g) and S. cerevisiae F-307 (64.86±4.47IU/gm wheat bran) but there was no significant difference between the last two microorganisms. The best microorganisms after that were A fumigates NRCF-113(60.24± 11.22IU/g); A.oryzae F -937(58.19± 2.67IU/g) and T. viride NRCF-107(54.41± 5.13IU/g) that there was no significant difference between them. According to the results, A. awamori NRC- F18 selected as the best fungus to complete all experiments for phytase production.
3.2.2. Evaluation of Different Wastes for Phytase Production
Screening of several wastes was involved. Phytase production varied with the type of wastes as shown in Table 3.
Presented results in Table 3 revealed that there was a significant difference between treatments and alkali treated rice straw was the best substrate for high phytase production reached 424.66± 2.92 IU /g rice straw followed by pomegranate peels)323.98±4.02IU/g pomegranate peels) then cotton seeds (133.75±1.35IU/g cotton seeds), wheat bran (106.26±0.89IU/g wheat bran),sugar bagasse pith (61.94± 1.98IU/g), orange peels (43.58±3.42 IU/g orange peels), then sorghum flour (38.30±0.38 IU/g) and cornflour (30.13±3.12 IU/g corn flour) but there was none significant difference between last two substrates. The last substrate after that was sugar beet pulp which gave lowest production (21.74±18.34 IU/g).
3.2.3. Effect of Nitrogen Source
Presented results in Table 4 revealed that there was a significant effect of nitrogen source in the growth medium of A. awamori NRC- F18 cultivated on alkali- treated rice straw under solid state fermentation. Ammonium phosphate (the main nitrogen source-control) and Ammonium acetate provide the highest phytase activity reached 258.75± 7.70 IU/g rice straw and 256.05±3.92 IU/g rice straw, respectively, but there was no significant difference between them. Then, there was a nonsignificant difference among ammonium oxalate)250.07± 3.91IU/g rice straw); ammonium nitrate (246.43±0.88IU/g rice straw); ammonium sulphate (245.46± 0.42IU/g rice straw); Potassium nitrate (240.91±5.86 IU/g rice straw; Diammonium hydrogen citrate (240.06±2.82 IU/g rice straw) nonsignificant difference between them. The best nitrogen source after that was urea (232.88± 8.63IU/g rice straw). The last substrate after that was ammonium hydrogen phosphate which gave the lowest production (153.01±2.54 IU/g).
| Fungus | Phytase Activity IU/g | Fungus | Phytase Activity IU/g |
|---|---|---|---|
| A.oryzae F -923 | 66.56 bc ±1.70 | A.fumigates NRCF-113 | 60.24 c ± 11.22 |
| A.oryzae F -937 | 58.19 c ± 2.67 | A. awamori NRC- F18 | 143.67 a ± 7.30 |
| A. niger F-258 | 110.62 b ± 9.85 | T.viride NRCF-107 | 54 .41 c ± 5.13 |
| S. cerevisiae F-307 | 64.86 bc ± 4.47 | ||
| LSD at α = 0.05 | 33.520 | ||
| Substrate | Phytase Activity IU/g | Substrate | Phytase Activity IU/g |
|---|---|---|---|
| orange peels, | 43.58 f ±3.42 | sugar beet pulp | 21.74 h ± 18.34 |
| pomegranate peels | 323.98 b ± 4.02 | cotton seeds | 133.75 c ± 1.35 |
| corn flour | 30.13 gh ± 3.12 | sorghum flour | 38.30 fg ±0.38 |
| sugar bagasse pith | 61.94 e ± 1.98 | wheat bran | 106.26 d ± 0.89 |
| rice straw | 424.66 a ± 2.92 | ||
| LSD at α = 0.05 | 12.1400 | ||
3.2.4. Effect of Carbon Source
The results presented in Table 5 showed that all carbon sources improved phytase production.
Glucose was the best carbon source for high phytase production reached 311.27± 8.65 IU /g rice straw rice straw then arabinose (289.02±18.59IU/g rice straw). They followed by fructose (277.79±6.04IU/g rice straw, dextran) (277.29 ±7.80IU/g rice straw), cellobiose (267.64±15.90IU/g rice straw), mannitol (264.88±4.59IU/g rice straw) and maltose (248.46±16.13IU/g rice straw). The rest carbon sources improved phytase production, but their effect was less than above-mentioned sources.
| Nitrogen Source | Phytase IU/g | Nitrogen Source | Phytase IU/g |
|---|---|---|---|
| Urea | 232.88 c ± 8.63 | Sodium nitrate | 233.41 c ± 22.68 |
| Ammonium hydrogen sulphate | 153.01d ±2.54 | Ammonium nitrate | 246.43 abc±0.88 |
| Diammoniumhydrogen citrate | 240.06 bc ±2.82 | Ammonium acetate | 256.05 a ± 3.92 |
| Ammonium oxalate | 250.07 ab ± 3.91 | Ammonium sulphate | 245.46 abc ± 0.42 |
| Potassium nitrate | 240.91 bc ± 5.86 | Ammonium phosphate | 258.75 a ±7,70 |
| LSD at α = 0.01 | 14.54 | ||
| Carbon Source | Phytase Activity IU/g | Carbon Source | Phytase Activity IU/g |
|---|---|---|---|
| Maltose | 248.46 de ± 16.13 | Glucose | 311.27 a ± 8.65 |
| Sorbose | 206.53 f ± 9.62 | Xylose | 229.64 e ±1.90 |
| Cellobiose | 267.64 cd ±15.90 | Lactose | 208.76 f ± 8.27 |
| Dextran | 277.29 bc ± 7.80 | Mannitol | 264.88 cd ± 4.59 |
| Galactose | 239.75 e ± 8.87 | Fructose | 277.79 bc ± 6.04 |
| Arabinose | 289.02 b ± 18.59 | Control | 267.75 cd ± 21.07 |
| LSD at α = 0.05 | 20.2900 | ||
3.2.5. Effect of Substrate% Flask Volume
The influence of substrate% on phytase production in the SSF was studied. Rice straw medium was prepared with different concentrations (1.5% to 5.5%). The results are shown in Table 6.
Presented results in Table 6 revealed that there was a significant effect on substrate concentration. Phytase production increased by increasing substrate concentration till 4.5% followed by decline when substrate concentration reached 5.5%. The best concentration for high phytase production was 4.5% reaching to 277.39± 12.05 IU /g rice straw and 4%)261.64±9.04IU/g rice straw but there was no significant difference between them and lastly it was 5%)256.72±5.23IU/g rice straw). This was followed by 5.5% (235.48±7.80IU/g rice straw); 3.5% (237.21±17.40IU/g rice straw) and 3% (231.27±5.95IU/g rice straw), but there was no significant difference between them. The lowest production observed with substrate concentration 1.5% reached 191.59 ±14.01IU/g rice straw.
|
Substrate: Flask Volume % |
Phytase Activity IU/g | Substrate: Flask Volume % | Phytase Activity IU/g |
|---|---|---|---|
| 1.5 | 191.59 e ± 14.01 | 4% | 261.64 ab ± 9.04 |
| 2 | 210.77 d ± 4.64 | 4.5% | 277.39 a ± 12.05 |
| 2.5 | 225.85 cd ±4.06 | 5% | 256.72 b ± 5.23 |
| 3 | 231.27 c ± 5.95 | 5.5% | 235.48 c ± 7.08 |
| 3.5 | 237.21 c ± 17.40 | ||
| LSD at α =0.05 | 17.0300 | ||
3.2.6. Effect ofIinitial pH
The effect of medium pH on phytase production was studied by adjusting the media pH between 3 and 7.5. The results are shown in Table 7.
Presented results in Table 7 revealed that there was a significant difference between treatments. The best pH for high phytase production was pH 6 and 5.5, which reached to 371.883± 0.822 IU /g rice straw and 363.184± 7.161 IU /g rice straw, respectively but there was none significant difference between them. This was followed by pH 5)307.349±6.625IU/g rice straw) and pH 4.5)300.011±1.755IU/g rice straw) but there was no significant difference between them. The rest of pH improved phytase production, but their effect was less than the effect of pH between (6 and 4.5). The lowest phytase production was at pH 3 reached 164.869± 7.77 IU /g.
| Initial pH Value |
Phytase Activity IU/g |
Initial pH Value |
Phytase Activity IU/g |
|---|---|---|---|
| 3.0 | 164.869 g ±7.78 | 5.5 | 363.184 a ± 7.16 |
| 3.5 | 194.048 f ± 2.84 | 6.0 | 371.883 a ± 0.82 |
| 4.0 | 231.771 e ± 1.17 | 6.5 | 275.771 c ± 6.39 |
| 4.5 | 300.011 b ± 1.76 | 7.0 | 256.352 d ± 8.17 |
| 5.0 | 307.349 b ± 6.63 | 7.5 | 193.941 f ± 1.55 |
| LSD atα=0.01 | 9.152 | ||
3.2.7. Effect of Moisture Level on Phytase Production
Influence of moisture level on phytase production in the SSF was studied and the results are shown in Table 8.
| Solid: Liquid Ratio |
Phytase Activity IU/g |
Solid: Liquid Ratio |
Phytase Activity IU/g |
|---|---|---|---|
| 1:1 | 214.549 e ± 3.12 | 1:4 | 371.883 a ± 0.82 |
| 1:2 | 282.379 d ± 1.16 | 1:5 | 327.723 c ± 11.34 |
| 1:3 | 349.483 b ± 8.67 | ||
| LSD at α = 0.01 | 11.98 | ||
The present work showed that there was a significant phytase production increase by increasing moisture level till 1:4 followed by a decline when moisture level reached 1:5. The maximum enzyme (371.883±0.82 IU /g rice straw) was observed when solid: moisture was 1:4. The lowest production observed with moisture level1:1 reached 214.549±3.12IU/g rice straw.
3.2.8. Effect of Inoculums Size on Phytase Production: Inoculum Concentration is an Important Factor for Microbial Growth and Enzyme Production
The results in Table 9 revealed that there was a significant difference between treatments and phytase production increase by increasing inoculums size till 2ml/5g followed by a decline when inoculums size reached 2.5ml/5g (391.723 ±1.793IU/gm). Inoculum concentration 2ml/5g provided the highest phytase production reacing 418.363± 16.709 IU/g. This was followed by inoculum concentration 1.5ml/5g (413.195 ±9.522IU/g), but there was no significant difference between inoculums size 1.5 and 2 ml/5g. The lowest production observed with inoculums size 0.5ml/5g reached 359.952 ±6.407IU/g.
3.2.9. Effect of Fermentation Period on Phytase Production
The influence of fermentation period on phytase production in the SSF was studied and the results are shown in Table 10.
Presented results in Table 10 revealed that there was a significant difference in enzyme production according to the incubation period. Phytase production increased by increasing the incubation period to 144 hr followed by decline when incubation period reached to 168 hrs. Incubation period for 144 hrs provided the highest phytase production reaching 437.195± 7.16 IU /g rice straw. This was followed by incubation period for 120 hrs)413.195±9.52IU/g rice straw) and phytase production decreased by decreasing incubation period and the lowest production after incubation for 24 hrs)24.928±3.91IU/g rice straw).
| Inoculum Size v/w(ml/5g) |
Phytase Activity IU/g Rice Straw |
Inoculum Size v/w(ml/5g) |
Phytase Activity IU/g Rice Straw |
|---|---|---|---|
| 0.5 | 359.95 c ± 6.41 | 2.0 | 418.36 a ± 16.71 |
| 1.0 | 371.88 c ± 0.82 | 2.5 | 391.72 b ± 1.79 |
| 1.5 | 413.19 a ± 9.52 | ||
| LSD at α = 0.01 | 16.57 | ||
3.2.10. Thermostability of Crude Enzyme
The effect of temperature on phytase activity was studied. Phytase assay was carried out at different temperatures and changes in the activity level are recorded (Table 11).
| Incubation Time (hr) |
Phytase Activity IU/g |
Incubation Time (hr) |
Phytase Activity IU/g |
|---|---|---|---|
| 24 | 24.928 f ± 3.91 | 120 | 413.195 b ± 9.52 |
| 48 | 101.728 e ± 1.90 | 144 | 437.195 a ± 7.16 |
| 72 | 285.195 d± 6.48 | 168 | 408.928 b ± 3 .91 |
| 96 | 388.128 c ± 2.23 | ||
| LSD at α = 0.01 | 9.886 | ||
The enzyme activity was checked at different temperatures. Statistical analysis represent that there was a significant difference between different temperatures and the most suitable temperature was 35°C phytase with an activity of 415.60 IU /gm followed by 40°C, activity was 413.97IU/gm); 45°C activity was)412.71IU/gm), 50°C activity was (400.88 IU/mg) and 55°C activity was)392.25 IU/gm) but there was no significant difference between them. It was found that, at 55°C, 60°C and 65°C, more than 90% of the activity remained and the percentage of decreasing in the activity was 5.62%, 5.99% and 6.35%, respectively. More than 80% of activity still remained at 70°C, 75°C and 80°C and the percentage of decreasing in the activity was 12.86%, 13.37% and 14.77%, respectively. At the same time, the enzyme lost 91.24% of its activity when the temperature shifted to100°C. According to the results, phytase was found to be thermostable. Maximum temperature at which phytase was active was 80°C.
| Temperature oC | Phytase activity IU/g | Relative activity % | % of Decreasing activity | |||
|---|---|---|---|---|---|---|
| 35°C | 415.60 | a | ± | 1.09 | 100 | |
| 45°C | 412.71 | abc | ± | 2.23 | 99.30 | 0.70 |
| 50°C | 400.88 | abc | ± | 30.40 | 96.46 | 3.54 |
| 55°C | 392.25 | abc | ± | 15.51 | 94.38 | 5.62 |
| 60°C | 390.72 | bc | ± | 1.10 | 94.01 | 5.99 |
| 65°C | 389.22 | c | ± | 7.65 | 93.65 | 6.35 |
| 70°C | 362.17 | d | ± | 6.24 | 87.14 | 12.86 |
| 75°C | 360.03 | d | ± | 12.52 | 86.63 | 13.37 |
| 80°C | 354.20 | d | ± | 21.74 | 85.23 | 14.77 |
| 100°C | 36.41 | e | ± | 10.39 | 9.86 | 91.24 |
| LSD at α = 0.05 | 24.360 | |||||
4. DISCUSSION
Multiple Sequence Alignments of phytases from genus Aspergillus were performed using CLUSTAL_O (1.2.4). Similar amino acids consisted of the same criterion placed in the same column. The first conserved amino acids glycine and proline in all selected organisms are at columns 43 and 59. These two amino acids often coincide with the extremities of well-structured beta strands or alpha-helices. Cysteines are famous for making C-C (disulphide) bridges. Conserved columns of cysteines usually indicate such bridges and provide a useful signature for recognizing protein domains and folds. The first conserved amino acids cysteines in all selected organisms are at position 77. Histidine and serine are often involved in catalytic sites, conserved histidine or a conserved serine are good candidates for being part of an active site. The first conserved histidine and serine in all selected organisms are at positions 88 and 30, respectively. The charged amino acids Lysine, Arginine, Aspartic Acid and Glutamic Acid, are often involved in legend binding. The presence of highly conserved columns of these charged amino acids indicates a salt bridge inside the core of the protein. Tryptophan is a large hydrophobic residue. It plays an important role in protein stability and is therefore difficult to mutate. When tryptophan mutates, it is usually replaced by another aromatic amino acid, such as phenylalanine or tyrosine. The first conserved amino acids tryptophan, phenylalanine, tyrosine in all selected organisms are at positions 307, 122 and 44, respectively. Patterns of conserved aromatic amino acids constitute the most common signatures for recognizing protein domains [40].
A. awamori NRC- F18 was selected as the best fungus for phytase production. This is supported by a study which concluded that A. awamori was the most suitable fungus for phytase production, as well as very important in the fermentation of such Japanese spirits and the most remarkable fungus applied for commercial phytase production [11, 12, 41-43].
The availability of the substrate material and its cost are the main factors for the selection of the ideal substrate for enzyme production in a solid-state fermentation process. This is related to dual role of solid materials, which supply nutrients in the fermentation medium and anchorage the fungus growth and enzyme release. Alkali treated rice straw was the best substrate for high phytase production. This is supported by several studies which concluded that rice straw is considered one of the abundant lignocellulosic wastes and sometimes creates environmental problems in some countries. Alkali treated rice straw is rich in cellulose and hemicelluloses as well as contains percentage of lignin and is familiar carbon source for lignocellulase production [26]. Different substrates were used for phytase production by fungi under solid state fermentation, such as wheat bran [9, 44]; rice bran; Canola meal; Groundnut cake; oilcakes; corn meal; rape seed; cassava bast and potato waste. Sporotrichum thermophile produced high phytase in SSF using sesame oil cake [16, 45-48].
Ammonium phosphate and ammonium acetate provide the highest phytase activity. The results are supported by several studies which concluded the significant role of nitrogen sources in fungal growth and phytase production [1, 28, 49].
Glucose was the best carbon source for high phytase production. This is supported by several studies which concluded that fungal strains prefer monosaccharids such as glucose and fructose for high production of enzyme that the fungus lack mechanisms for converting polysaccharides into simpler glucose molecules. According to the present study, glucose is considered the best carbon source for industrial fermentations [29].
Phytase production increases by increasing substrate concentration followed by decline. The results supported by several studies concluded that increasing of rice bran concentration above 1% resulted in inhibition of phytase activity due to more inorganic phosphate in the medium while decreasing of rice bran concentration resulted in less phytase activity due to insufficient inorganic phosphate in the medium [30].
Microbial production of enzyme phytase depends on the extracellular pH. Most enzymatic processes and transport of various components across the cell membranes are influenced by culture pH [6, 50]. The best pH for high phytase production were pH 6 and 5.5. This was supported by several studies which concluded that phytase has pH optimum range (pH 4.5-6.0) with a rapid drop-in activity at pH value above 6.0 [38, 51]. Similar results were reported by a study which stated that production of phytase from Sporotrichum thermophile was maximum at pH 5 [52]. Phytase production has been mostly reported to be acidic with a neutral pH range. While other studies found that Arxula adeninivorans showed maximum activity at pH 5.5 and A.oryzae produced the highest phytase titers at pH 6.5 [1].
Moisture level content is a critical parameter for fungal biomass production and enzyme biosyntheses in solid-state fermentation [26, 53]. The microbial growth and product formation occur near the surface of the solid substrate particle having low moisture contents [26]. The optimum level of moisture varies according to substrates, size and microorganisms [54]. Phytase production increased by increasing moisture level till 1:4 followed by a decline when moisture level reached 1:5. The results are supported by several studies which concluded that moisture cause swelling of the substrate and facilitates utilization of the nutrients present in the medium by the fungus. The solubility of the nutrients in the substrate decreased at low moisture level. While higher moisture content resulted in a reduction in the substrate porosity, it decreased the availability of oxygen for fungus. Consequently, oxygen transfer affects nutrient metabolism, causing inhibition for enzyme biosynthesis. Other studies found a relation between moisture level and nature of the substrate to permit suitable growth and enzyme secretion in fermentation medium by specific microorganism. Moisture content was studied extensively by researchers working in solid state fermentation systems and found that fungus can biosynthesize optimum enzymes activity under a range of moisture content or at a limited moisture level. A. carbonarius produced optimum phytase when moisture percentage ranging from 53 to 60% cultivated on canola meal. While 60% was reported for penecillium spp. when cultivated on fine particles of crushed fava bean [31].
Inoculum concentration is an important factor for microbial growth and enzyme production. Phytase production increases by increasing inoculums size till 2ml/5g followed by a decline when inoculums size reached 2.5ml/5g. The results were supported by several studies which concluded that the enzyme synthesis increase with increase in inoculums concentrations followed by steady decline with increasing inoculum concentrations. This is due to an increase in the fungal biomass production, which results in increased competition for nutrients, exhaustion and imbalance of nutrients which results in reduced enzyme production. Similar findings were reported with Mucor racemosus [1, 33]. Phytase production increased by increasing incubation period till 144 hr followed by a decline when incubation period reached 168 hrs. The results were supported by several studies which concluded that the enzyme production increased with incubation time. The enzyme yield declined during further incubation due to the reduced nutrient level of the medium. A similar finding in the production of phytase was reported with Mucor racemosus. Another study reported that highest phytase production by A. niger and A. tubingensis were after 4 days [1, 33]. Phytase was found to be thermostable. Maximum temperature at which phytase was still active was 80°C. The results were supported by several studies, which concluded that phytase enzyme was thermostable and 80% of the activity still remained when the temperature was shifted to 70°C [11, 38, 55, 56s].
CONCLUSION
Bioinformatic studies showed multiple alignments between the amino acid sequences of phytases for Aspergillus spp. enabled the identification of a highly conserved sequence. The results revealed that there are 88 identical positions and 135 similar positions between these sequences but the identity percentage is 16.858%. These results explain the difference in activity of phytases produced from different species although they have the same function. These results were followed by experimental validation of phytase production by some microorganisms. The experimental study has allowed the screening of seven phytase producing microorganisms namely A. oryzae F -923; A. oryzae F -937; A. niger F-258; A. awamori NRC- F18.; Aspergillus fumigates NRCF-113 and T.viride NRCF-107 as well as S. cerevisiae F-307. A.awamori NRC- F18, which provided the highest phytase activity selected as the best fungus to complete all experiments through solid state fermentation which proved as a good method for enhancing phytase production. Present study on phytase production under SSF by A. awamori NRC- F18 indicates that pretreatment of agriculture residues such as rice straw was useful for the production of phytase by utilization of less expensive substrate. Enrichment of rice straw with glucose, Ammonium phosphate as carbon and nitrogen sources at pH 5.5, temperature 30°C resulted in better yield of enzyme. The optimization experiments showed that a maximum phytase production obtained by A. awamori NRC- F18 using rice straw as a waste, incubation time was 144 h. The maximum enzyme was observed when solid: moisture was 1:4. phytase was found to be thermostable. Maximum temperature at which phytase was still active was 80°C. The present study suggests that A. awamori NRC- F18 strain has a significant value for production of phytase and it can be used in industrial production of phytase. Supplementation of animal feed with phytase has an important role in improving the nutritional status of feed and improving digestion in mono gastric animals.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
Not applicable.
HUMAN AND ANIMAL RIGHTS
Not applicable.
CONSENT FOR PUBLICATION
Not applicable.
AVAILABILITY OF DATA AND MATERIALS
The data supporting the findings of this study are present within the article.
FUNDING
None.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
The instruments and needed materials were supplied by Microbial Chemistry Department, Genetic Engineering and Biotechnology Division, National Research Centre, Dokki, Giza, Egypt.

