All published articles of this journal are available on ScienceDirect.

Artemia, Probiotics, And Immunostimulant: Essential Elements For Optimal Litopenaeus vannamei Shrimp Production
Abstract
Aquaculture is a vital component of the global food supply chain. Crustacean farming, primarily focused on shrimp species, is a vital component of aquaculture. Currently, the annual production of shrimp is approximately five million metric tons, and it is expected to increase to meet the global demand. Feed is the single most crucial factor for culturing shrimp, accounting for more than half of the input cost. Several feeds, including algae, zooplankton, basal feed, plant-based feed, and small mollusks, have been used to feed the larvae and grow shrimp. However, Artemia/Brine shrimp is the most commonly used feed given to shrimp during their early stages. There is no commercial feed that can fully replace artemia, as it meets the nutritional requirements, has better digestibility, and is convenient to prepare. Similarly, an intensified shrimp farming system aimed at increasing production has rendered shrimp more vulnerable to pathogens and diseases. Antibiotics have been used in shrimp farming, but it has adverse effects in the long run. Hence, probiotics and immunostimulants have been used as feed to enhance the immunity and overall health of shrimp, thereby improving production. Probiotics are live organisms that improve the host’s health by modulating the gut microbiome. Immunostimulants, on the other hand, improve the immune system of shrimp by directly interacting with its innate immunity system. This paper discusses the benefits and functions of artemia, probiotics, and immunostimulants on shrimp health and overall production. All these food sources play a crucial role in shrimp growth and production when used effectively.
1. INTRODUCTION
In recent years, the total aquaculture production has surpassed 700 tons [1]. With this, the aquaculture industry has become an integral and essential part of the global food chain, supplying the global population with affordable, high-quality aquatic food for consumption. It is expected to remain so for years to come [2]. Currently, there is an ongoing debate regarding food security and feeding a global population of over 8 billion [3]. Due to the growing population, aquaculture is expected to evolve to meet the increasing demand for protein derived from aquatic sources [4]. Shrimp production accounts for more than 50% of the total crustacean production, which in turn accounts for 23% of the global aquaculture production [1]. Hence, shrimp farming has become one of the most widely traded seafood products in the world and has become a fundamental part of aquaculture [1]. The shrimp industry has experienced rapid development in recent years to meet the increasing global demand for seafood [5, 6]. Since capture fisheries are unable to meet the rising demand, the shrimp industry is gradually taking over [7]. Due to its economic significance, the shrimp industry has been recognized as one of the aquaculture systems with the fastest growth rate in recent decades [8]. Feed accounts for up to 50% of the total cost of shrimp production, playing a significant role in the industry's economic aspect [9]. Determining the dietary requirements of a cultured species is a crucial component of farming [10]. Different dietary supplements are used in shrimp culture to enhance the immune system, promote body growth, improve nutrient absorption, and boost the function of digestive enzymes in shrimp [11]. A proper feeding regimen guarantees the growth of the cultivated species to its full potential with a minimum amount of feed waste [12, 13]. Hence, an important domain of research is developing a proper and balanced feed, along with a quality dietary supplement to meet its nutritional demands and ensure its better growth [10, 14].
The primary food source for larvae culture is live feed, which is especially crucial when raising marine species larvae. Aquaculture species in their early state are still fed live feed [15]. Artemia nauplii is the most widely utilized live feed for rearing larvae in aquaculture industries [16] due to its ideal size for the larvae, high nutritional value, and ease of digestion. Artemia is the primary meal for shrimp larvae [17]. It is assumed that even in the future, it won’t be advisable to substitute live feeds with any other sources, such as commercial feed or fish meal, because of their movement that triggers the hunting instinct in larvae [18]. Other methods of feeding are also being opted for to improve shrimp production [19]. For instance, probiotics, which are live microorganisms that are non-pathogenic in nature and reside in the intestine or gut of the host and improve their health [20]. This is achieved by enhancing the immune system [21]. When added to the water system, it can also enhance water quality, thereby keeping harmful bacteria in check and preventing disease [22]. Since vertebrates like shrimp rely on nonspecific immune function, probiotics can act on these systems and influence them to fight against pathogens [23]. Similarly, immunostimulants are natural occurring substances that can also be used to prevent disease and enhance the immune system [24, 25]. The process by which these natural additives work against bacterial resistance is by modulating the gut microbiome/microbiota and enhancing immune functions, or directly activating innate systems in shrimp [26]. The proper use of immunostimulants can help in maintaining sustainability in shrimp aquaculture [3]. This paper will discuss the benefits of artemia, probiotics, and immunostimulants on shrimp aquaculture.
2. METHODOLOGY
To develop this review, an extensive literature search was conducted to gather and synthesize information related to the use of Artemia, probiotics, and immunostimulants in the culture of Litopenaeus vannamei. The goal was to compile current findings, summarize experimental outcomes, and identify emerging practices in shrimp aquaculture.
Relevant scientific literature was gathered using multiple online databases, including PubMed, Science Direct, Scopus, Web of Science, and Google Scholar. The search focused on peer-reviewed articles, review papers, and official reports, primarily published between 2000 and 2024, although earlier landmark studies were also included where necessary. Keywords such as “Artemia,” “shrimp live feed,” “Litopenaeus vannamei,” “probiotics in aquaculture,” and “shrimp immunostimulants” were used in various combinations to capture a broad spectrum of research related to shrimp nutrition, health, and survival.
After retrieving the initial body of literature, articles were carefully screened for relevance. Studies were selected based on their focus on L. vannamei or related shrimp species and their contributions to understanding the nutritional and immunological impact of artemia, probiotics, or immunostimulants. Articles that lacked methodological clarity or focused on unrelated aquatic species were excluded. Ultimately, more than 100 relevant papers were reviewed in detail and used to inform the content of this manuscript.
The selected studies were read thoroughly and analyzed to extract key information. These included the biological role and enrichment potential of Artemia nauplii as a live feed, the mechanisms by which probiotics enhance gut health and disease resistance in shrimp, and the immune-boosting properties of various natural immunostimulants. Information from laboratory experiments, field studies, hatchery trials, and meta-analyses was integrated to provide a comprehensive overview.
In cases where comparisons were available, findings were organized to highlight differences in performance among various strains of probiotics or enrichment methods used for artemia. Several figures and tables were developed to summarize these comparative findings and mechanisms of action. Attention was also paid to gaps in current research, such as the long-term effects of immunostimulants or the challenges in replacing live feed with commercial diets.
2.1. Importance of Artemia
The primary food source for culturing larvae in aquaculture is live feed, which is especially crucial when raising larvae of marine species [15, 27]. A sufficient supply of nutrient-rich live food remains a critical component for successful larval rearing, and zooplankton live food is the fundamental building block of larviculture [28]. Among the various live feeds available, Artemia and rotifers are the preferred live feeds [18]. Artemia belongs to the phylum Arthropoda and has appendages that are shaped like leaves arising from its appendages [29]. Its ability to survive in hypersaline environments worldwide makes it distinct [30]. Over the past 50 years, the Great Salt Lake (GSL) has been the main source of Artemia cysts [31]. It is readily available and can be quickly hatched in large quantities at any time [32]. Artemia franciscana and Artemia salina are commonly used species of Artemia [33]. Because they hatch easily, Artemia nauplii are the most practical live feeds for aquaculture [34]. They have evolved to survive in harsh environments that include high temperatures, low oxygen concentrations, desiccation, complex ionic composition (water rich in sulfate, chloride, or carbonate), and intense UV radiation [35]. When a newborn is first fed, its digestive system is still very basic, and its stomach is lacking [36]. Especially the altricial larvae (immature and underdeveloped larvae) lack an intestine, and epithelial cells in the hindgut are responsible for protein absorption [31]. Such a digestive system is unable to process prepared diets in a manner that allows the larvae to grow and survive in a manner similar to those fed live feeds [37, 38]. Artemia nauplii have been reported to be highly digestible when fed to aquatic species, and their Protein Efficiency Ratio and Net Protein Utilization are elevated [39]. Artemia has the presence of digestive enzymes like amylase and trypsin in it [40], and these enzymes might digest Artemia's own tissue, causing self-digestion and making it easier for larvae to digest it [34]. Moreover, because of their movement in the water column, larvae can easily spot and prey on them. In contrast, formulated or commercial diets tend to settle quickly on the bottom or at the water surface, within minutes, and thus become less accessible to the larvae [15]. Its vivid body color also makes it quite discernible [34]. The larvae's diet and nourishment are among the most important factors that restrict the production of shrimp juveniles. Artemia, also known as Artemia supplementation, is usually given during the mysis stage in commercial larviculture of penaeid shrimp, and the initial feeding starts with live microalgae [41]. Artemia constitutes 40-67% of protein, and the percentage varies according to the different life stages of artemia [42]. Instar-I-staged Artemia nauplii molt into Instar-II within a period of 6 to 8 hours. Usually, Instar-I is preferred because it is smaller, more perceptible, and quicker to swim, making it harder to capture [43]. They also have higher fatty acid and energy content [34]. These nauplii can also be stored in cold conditions for a period, causing them to become inactive, which will be beneficial for shrimp species like black tiger shrimp to capture [44]. When Artemia was used during larval rearing, many species, including shrimp, have shown decreased mortality and increased growth rates.
2.2. Artemia Enrichment
However, one drawback these nauplii have is the absence of polyunsaturated fatty acids (PUFAs), specifically Eicosapentaenoic acid (EPA) and Docosahexaenoic acid (DHA) [45]. Hence, to fill in the nutritional gap of artemia, enrichment must be done [33, 31]. Exposing nauplii to specific substances high in HUFA content can result in enhanced Artemia nutritional value, improving growth, survival, and net production of both finfish and shellfish larvae [33]. Nauplii can filter, ingest, and absorb these substances, and when consumed by larvae, can readily deliver them into the larvae [33]. In embryos and larvae, fatty acids are the major source of energy; however, they are unable to produce it on their own, and hence, artemia must be enriched with fatty acids to fulfill this requirement [46]. Based on the species and nutritional requirements, the rate of enrichment can be altered [33]. Because of its importance in fish and shrimp larvae culture, HUFA is the most studied nutrient in artemia [47], and numerous studies have been conducted to investigate the impact of enriched artemia on these larvae [33]. Farfantepenaeus paulensis, also known as the pink shrimp, that fed upon HUFA-enriched Artemia, exhibited increased endurance to factors such as ammonia and salinity stress, and showed better weight gain compared to shrimp fed unenriched Artemia [48]. The tissues of the PLs were also found to have greater EPA and DHA content as compared to the control group. Both DHA and EPA play a crucial role in the regular function of cells, such as osmoregulation, formation of membranes, and immunity [49]. HUFA-enriched artemia not only increased DHA and Arachidonic Acid (ARA) levels in white leg shrimp but also showed healthier hepatopancreas with healthy and larger B cells and less damaged tissue [50]. Similar results were found in studies involving other species, such as the Asian sea bass, where enrichment resulted in better weight gain and fewer skeletal deformities [51]. Likewise, Pacific cod larvae feeding on HUFA-enriched Artemia showed better resistance to an air exposure test [52]. Because of the low amounts of DHA and EPA, unenriched artemia can be insufficient for marine larvae [46, 53]. However, freshwater fish are known to undergo various enzymatic processes, such as desaturation and elongation, to produce Long Chain Polyunsaturated Fatty Acids (LCPUFAs), including DHA, EPA, and ARA [54, 55]. In another study, Litopenaeus vannamei post-larvae (PL) showed better growth and survival rates when given different levels of artemia enriched with PUFA (1, 2, and 3%) compared to PLs given unenriched artemia [56]. Likewise, artemia enriched with gamat emulsion also significantly improved weight and survival rates in Pacific white leg shrimp [57]. Higher content of gamat emulsion boosts the growth rate. Gamat emulsion can be obtained from sea cucumber [57] and is a rich source of protein and DHA [58]. However, very limited research has been done to use gametemulsion as an enriching factor for artemia. Besides fatty acids, artemia also lacks various essential nutrients required by marine larvae; hence, different methods have been established to enrich artemia with these essential nutrients [59]. Elements like minerals such as Selenium [59] and amino acids like Methionine [60] have been used as an enriching medium for Artemia. Similarly, vitamins like C and E have also been used for Artemia bioencapsulation to enhance the growth of fish larvae [61, 62]. The death rate of marine larvae may surge due to a shortage of these essential nutrients [63]. Microalgae are also used as an enriching medium. Artemia enriched with Chaetoceron calcitrans showed a higher amount of essential fatty acid content than the unenriched ones and increased specific growth rate in Macrobrachium americanum [64]. Hence, enrichment causes beneficial effects on animals.
The increased demand for shrimp is met by adopting intensive/super-intensive farming; however, this type of farming has several impacts, including deteriorating water quality, excessive waste, and other environmental problems [65]. This will increase the risk of disease outbreaks and mortality in shrimp [66]. Antibiotics/ chemotherapeutics are commonly used for
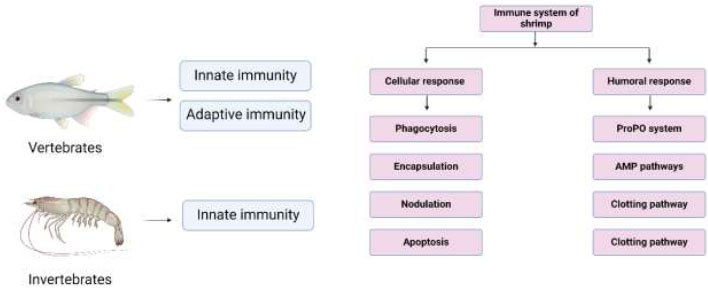
Comparison of immune systems in vertebrates and invertebrates, and a schematic overview of shrimp immunity.
disease mitigation in shrimp aquaculture [67]; however, these elements have a negative impact on the microflora, causing harmful substances to accumulate and creating bacteria that are drug-resistant [68]. Studies have shown that probiotics can improve the immunity of a host and enhance its growth by preventing the growth of harmful pathogens [69, 70]. Fuller (1989) [71] defined them as live microorganisms that promote a healthier gut microbiome and improve host health. Probiotics can perform multiple functions, such as colonization of the gut, exhibit antagonistic activity, facilitate digestive function, and produce beneficial nutrients like biotin, while also facilitating waste excretion [72]. Because invertebrates have a non-specific immune system that protects them from pathogenic organisms, probiotics have been shown to influence this system and produce transduction signaling molecules, which can alert the immune system to defend against pathogenic bacteria [23]. Similarly, immunostimulants are natural occurring substances that can also be used to prevent disease and enhance the immune system [24, 25]. The process by which these natural additives work to resist bacteria is by modulating the gut microbiome/microbiota and enhancing immune functions, or directly activating innate systems in shrimp [73]. The proper use of immunostimulants can help in maintaining sustainability in shrimp aquaculture [3]. Shrimp have a robust innate immunity system but lack an adaptive immunity system [3]. An overview of the shrimp immunity system is shown in Fig. (1).
2.3. Probiotics
Shrimp are highly dependent on their culture conditions, and alterations to this system can cause an increase in pathogenic load; hence, meticulous strategies must be adopted to prevent this [74]. Probiotics perform multiple functions in hosts by inhibiting the formation of bacterial or pathogenic colonies, enhancing enzymatic actions, and increasing the growth rate [75]. The primary action performed by Probiotics (also known as non-pathogenic bacteria) is to alter host-related microorganisms [76, 77]. Since gut space, materials, and resources are limited, colonization of good bacteria can impede the growth of pathogenic bacteria by competing for resources and space [78]. These bacteria then produce substances, such as antimicrobial compounds, that put unwanted invaders or pathogens in check [79]. When added to the water system, it can also enhance water quality, thereby controlling harmful bacteria and preventing disease [22]. Because invertebrates have a non-specific immune system that protects them from pathogenic organisms, probiotics have been shown to influence this system and produce transduction signaling molecules, which can alert the immune system to defend against pathogenic bacteria [23]. Probiotics are involved in various functions, including colonization of gut microorganisms, exhibiting antagonistic action towards certain substances, enhancing digestive function, and producing beneficial nutrients such as biotin, as well as facilitating waste excretion [72]. Once probiotics are taken, they increase Short Chain Fatty Acids (SCFA), which initiates insulin-like growth factor 1 (IGF-1) production [80]. IGF-1 then binds cell surface receptors and induces cell differentiation and proliferation [81]. SCFA plays an important role in modulating the immune system, improving host defenses, and increasing signaling molecules for gene expression [82]. It also enhances endogenous enzymes that improve digestion and absorption, thereby enhancing gut health [82]. Shrimps that were fed probiotics have shown an increased height of microvilli in their gut, which significantly increases nutritional absorption in the gut [83, 84]. Probiotics contribute to overall immune system enhancement, which also promotes improved growth [84]. When beneficial bacteria are more abundant in the gut, the performance of shrimp is better. However, if harmful bacteria are more numerous, shrimp are more likely to contract disease and exhibit reduced growth [19]. Probiotics also enhance carbohydrate hydrolysis and amylase activity [85] and support the growth of beneficial gut bacteria, such as Lactobacillus and Bifidobacterium, in the gut [86]. There are only a few studies that have shown how probiotics function in a host, and the action mechanism of probiotics in aquaculture is not well studied [74]. Fig. (2) illustrates the proposed functional mechanism by which probiotics exert beneficial effects in shrimp.
The comparative benefits of various probiotic strains on shrimp growth and immunity are summarized in Table 1.
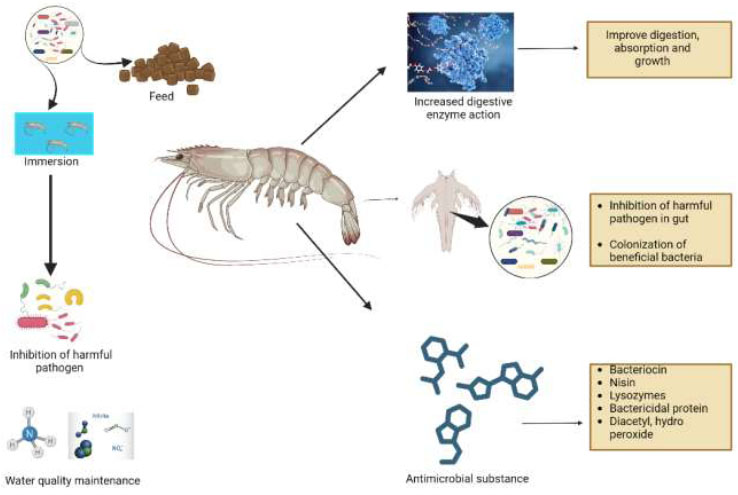
Functional mechanisms of probiotics in shrimp aquaculture.
| Probiotics | Treatments | Shrimp species | Benefits | Citations |
|---|---|---|---|---|
| Photosynthetic bacteria and Bacillus spp. | T1-1 g/kg Photosynthetic bacteria + 1 g/kg Bacillus spp T2-5 g/kg Photosynthetic bacteria + 5 g/kg Bacillus spp T3-10 g/kg Photosynthetic bacteria + 10 g/kg Bacillus spp Control |
Litopenaeus vannamei | Enhanced growth, protease, and amylase activity as compared to the control group. | [87] |
| Lactobacillus acidophilus, Rhodopseudomonas palustris, and Bacillus coagulans | T1-Lacidophilus T2-Rhodopseudomonas palustris T3-Bacillus coagulans |
Litopenaeus vannamei | Better growth performance and immune response as compared to the control. Shrimp given Bacillus showed the highest Phenoloxidase and Superoxide dismutase activity. |
[88] |
| Bacillus spp. | C: Control T1: Bacillus sprayed live. T2: Bacillus in frozen dry form |
Penaeus monodon | Better growth performance and feeding efficiency. Vibrio’s population was reduced in both the culture system and in the gut and hepatopancreas. |
[89] |
| Lactobacillus, Bacillus, Nitosomonas and Nitobacter | C: Control T1: 1 ml/10 L T2: 2 ml/10L T3: 3 ml/10 L T4: 4 ml/10 L C: Control |
Litopenaeus vannamei | Better results in terms of weight gain, yield, and feed efficiency were seen in probiotic-treated shrimp, especially in T2. | [90] |
| L. fermentum, L. pentosus, Saccharomyces cerevisiae, and Bacillus subtilis | LW2: L. fermentum BD6: L. pentosus P13: Saccharomyces cerevisiae E20: Bacillus subtilis MD-Mixture |
Litopenaeus vannamei | Mixture group enhanced survival, resistance against Vibrio, activity of immune genes PO, lysozyme, and release of reactive oxygen species. | [91] |
| Mixed probiotics | T1: 0 mg/kg feed T2: 2000mg/kg T3: 4000 mg/kg T4: 6000 mg/kg T5: 8000 mg/kg |
Litopenaeus vannamei | Probiotics enhanced growth performance, activities of SOD, catalase, more variability in microbiome population, as well as increased villi of the intestine’s length. | [92] |
| Bacillus subtilis and B. licheniformis | T1: 2 g/100 g feed T2: 4 g/100 g feed T3: 6 g/100 g feed T4: 8 g/100 g feed T5: 10 g/100 g feed |
Penaeus monodon | The results showed improved growth rate and activity of enzymes involved in digestion with an increase in the amount of probiotics in the diet. The activity of protease, amylase, and lipase was increased. | [93] |
| Bacillus subtilis | T1: 102 CFU/ml T2: 103 CFU/ml T3: 104 CFU/ml T4: 105 CFU/ml C: Control |
Litopenaeus vannamei | Probiotics resulted in improved growth and immunity by enhancing phagocytic activity, PO, lusozyme, and Alf action, as well as increasing microvilli length and the width of the intestinal wall. | [94] |
2.3.1. Immunostimulants
An immunostimulant is a natural substance that enhances the host's immune system [25]. It is a good choice for managing shrimp immunity and health [3]. In aquaculture, its application has started recently; however, its development is happening at a rapid pace, with the utilization of different types available [95]. When inside the body, it stimulates macronutrients, nanoparticles like chitosan, and other elements [96]. This causes an enhanced humoral and cellular response in various parts of the body, such as the blood, head kidney, and immune genes are upregulated [97, 98]. Beta-glucans are an immunostimulant that have demonstrated a 0.1% increase in intestinal microbial richness of L. vannamei, upregulation of immune genes, and activation of ACP (Acid phosphatase) and AKP (Alkaline Phosphatase). ACP and AKP are important for enhancing and facilitating phagocytosis, a crucial process in defense [99]. Similarly, prebiotics as immunostimulants in the gut can be metabolized by Probiotics like Lactobacillus and Bifidobacterium to generate Short Chain Fatty Acid (SCFA) and lactate [75]. SCFA prevents the increase of harmful bacteria by decreasing stomach pH and stimulates the growth of beneficial bacteria, such as LAB and Bifidobacterium [100]. Immunostimulants have been shown to enhance phagocytosis activity, respiratory burst, and SOD activities, and prevent pathogenic diseases in white leg shrimp [101]. In another study, the use of immunostimulants resulted in a reduction of Vibrio in shrimp hepatopancreas, and the actions of phagocytes, PO, and SOD were greatly enhanced compared to the control group [102]. Marine algae-derived immunostimulants have demonstrated resistance in whiteleg shrimp when challenged with White Spot Syndrome Virus (WSSV) by improving Haematocyte count, granule cell number, and respiratory bursting [103]. Genipin (GN) is bioactive in nature and is extracted from Gardenia jasminoides [104]. When administered at a rate of 50 mg/kg per day, it was observed that GN slows down WSSV multiplication and impedes Bax inhibitor-1 gene expression, resulting in further retardation of the WSSV infection rate in shrimp. It is used as an effective method to prevent WSSV in shrimp [104]. Immunostimulants derived from different herbal plants, such as Cyanodon dactylon, Tinospora cordifolia, and Picrorhiza kurrooa, have been shown to decrease viral load and increase survival (74%) in shrimp [105]. Cathelicidin-5 can be extracted from Alligator sinensis and utilized to cure WSSV in shrimp. Exopalaemon modestus was injected with WSSV and fed cathelicidin, showing reduced mortality, lower viral protein (VP28) multiplication, and enhanced activity of antioxidant enzymes and immune genes [89]. Hence, immunostimulants are used in shrimp culture to enhance the immunity and health of shrimp by improving gut health, augmenting immune response, and increasing survival.
The outermost exoskeleton in shrimp serves as the first line of defense against pathogens and foreign elements, thereby preventing them from entering the body [106]. Once they move past the outer layer, the pathogen reaches the hemocoel in shrimp, which consists of hematocytes (chief cells in shrimp immunity) [107]. Hematocytes detect these pathogens through the presence of Pathogen-Associated Molecular Patterns (PAMPs). PAMPs are the molecular patterns distinctive to pathogens, based on which hematocytes, which detect non-self cells, recognize molecules such as Peptidoglycan (PG) and Lipopolysaccharide (LPS) [108]. Hematocytes can detect and distinguish between foreign cells and self-cells with the aid of receptors known as Pattern Recognition Receptors (PRRs). Once PRR can distinguish between these two cells, an activation of the cellular signaling cascade results in both cellular and humoral responses [109]. PRR also identifies the PAMP of immunostimulants, and instead of causing a targeted response to a specific pathogen, it elicits a broad immune response that helps shrimp identify and fight against pathogens more efficiently [3, 110]. As illustrated in Fig. (3), immunostimulants enhance the shrimp's immune system by activating multiple defense pathways.
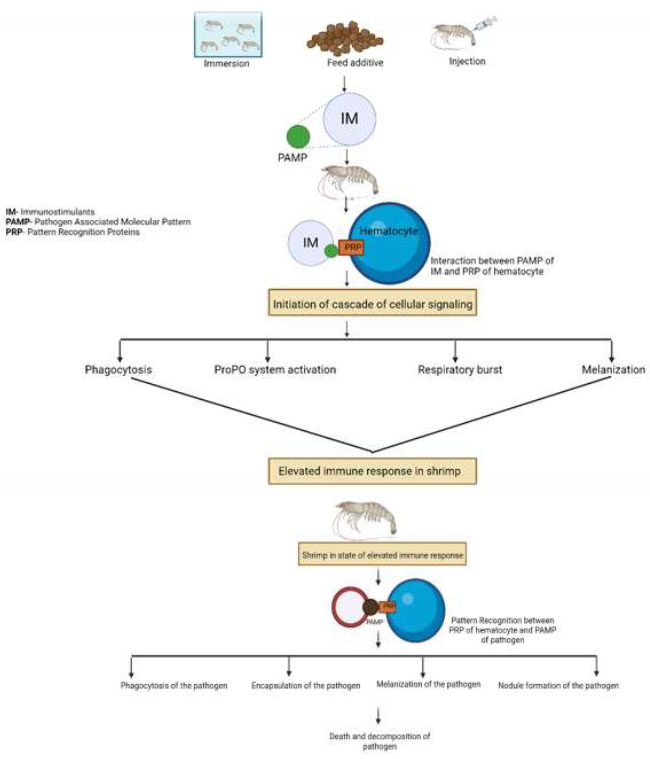
Schematic representation of immunostimulant action in shrimp immunity.
3. DISCUSSION
Due to its convenience [32], hatchability [34], and easy detectability by larvae [15], artemia is one of the most preferred live feeds. Since it lacks fatty acids like EPA and DHA [45], bio-enrichment with fatty acids are carried out [46]. However, nauplii gut can only hold a limited amount of nutrients, and enhancement of one nutrient can cause a reduction of another element, such as microminerals [59]. Efforts have been made to develop a feed for larvae; at present, it is believed that no commercial feed can fulfil the demand of crustacea as well as fish larvae [28]. Though widely recognized as an excellent choice of live feed for shrimp and fish larvae, Artemia nauplii’s unhatched cysts and empty shells can pose a threat in aquaculture hatcheries. The cysts are indigestible and could lead to intestinal blockage or other negative effects when consumed by the larvae [111-114]. Decapsulation is a chemical process used for the complete removal of the hard outer shells of cysts, also known as the alveolar layer [115]. This process not only reduces the chances of gut obstruction due to cyst shell ingestion by the larva but also improves the hatching rate, the nauplii’s energy content, and disinfects the cysts [116]. Similarly, SEP-Art Technology provides magnetic Artemia eggs that ensure the highest performance. By creating a special coating on the cysts, D-Fense inhibits the growth of harmful pathogens.
Although Artemia does not act as a vector for pathogens [117], careful handling and proper enrichment practices remain essential to minimize potential disease risks and enhance shrimp survival [118]. Probiotics are used as disinfectants; however, studies are needed to identify pathogens in artemia [119]. Probiotics produce chemical substances that create an environment unfavorable to harmful pathogens, preventing their multiplication and spread [120]. Probiotics perform multiple beneficial functions in the host [72]. However, they develop resistance to antibiotics, and hence, in the long run, their application is called into question [16, 110]. Similarly, the probiotics used need to be of higher quality and appropriate; otherwise, negative results may occur [121]. Probiotics manufacturing requires modern and challenging techniques [87]. Another limitation it might have is a slow response, i.e., it works more slowly compared to antibiotics [121].
Immunostimulants primarily enhance immunity in shrimp against pathogens through the activity of phagocytosis cells; however, some bacteria are immune to phagocytosis and can persist within macrophages, such as Renibacterium and Pseudomonas, among others. In these cases, immunostimulants are ineffective [98, 121]. Studies have shown that immunostimulants induce pathogen resistance, growth, and enhanced immunity; however, these effects are mainly short-lived, and very few studies have been conducted to determine their long-term effects. The long-term effect of immunostimulants is unknown. Some studies have shown it to be effective, whereas others have not [67]. Immunostimulants may have a negative impact by overstimulating the immune system. It might cause hemocytes to overreact, leading to an excessive inflammatory response and the rapid release of harmful substances, which can have a negative impact on its important cells. This can cause hemocytes to clump, reducing the number of free hematocytes available and ultimately causing the immune system to exhaust itself [14, 67]. While immunostimulants can enhance shrimp immunity, they must be used with caution. Overstimulation of the immune system can lead to a range of adverse effects, including tissue damage, reduced availability of immune cells, toxicity, depletion of immune resources, and increased energy expenditure. These potential drawbacks underscore the importance of using immunostimulants in shrimp aquaculture in a balanced and controlled manner.
CONCLUSION
In this paper, the roles of Artemia, probiotics, and immunostimulants in Litopenaeus vannamei shrimp culture are investigated, highlighting their impact on shrimp growth, immune function, and overall health. Due to its suitable nutritional composition and easy availability, artemia remains the superior live larval feed. However, its enrichment is essential to meet all dietary requirements. Probiotics and immunostimulants, on the other hand, can become a potential substitute for antibiotics in sustainable shrimp farming, as they boost immune responses and support the gut health of the host. Overall, our results highlight the importance of combining suitable live feeds and nutritional supplements for shrimp production enhancement. For a better understanding of their long-term impacts and optimal applications in shrimp aquaculture, further investigation and studies in commercial farming settings for an extended period are required.
AUTHORS' CONTRIBUTIONS
The authors confirm their contributions to the paper as follows: study conception and design: YR; Writing - Reviewing and Editing: AS. All authors reviewed the results and approved the final version of the manuscript.
LIST OF ABBREVIATIONS
| CFU | = Colony Forming Unit |
| ml | = Milliliters |
| G | = Gram |
| kg | = Kilograms |
| PO | = Prophenoloxidase |
| SOP | = Superoxide Dismutase |
| Alf | = Antilipopolysaccharide Factor |
| T1, T2, etc. | = Treatment 1, Treatment 2, etc. |
| SCFA | = Short-Chain Fatty Acids |
| IM | = Immunostimulants |
| PRP | = Pathogen Recognition Proteins |
| PAMP | = Pathogen-Associated Molecular Pattern |
AVAILABILITY OF DATA AND MATERIALS
The data supporting the findings of the article is available in the Zenodo repository at https://doi.org/10.5281/zenodo.16933014, reference number 10.5281/zenodo.16933014.
ACKNOWLEDGEMENTS
None declared.

