All published articles of this journal are available on ScienceDirect.

Molecular Study of Klebsiella Oxytoca Associated with Urinary Tract Infection in Children
Authors Info & Affiliations
Abstract
Background:
There are scarce reports about the association of Klebsiella oxytoca (K. oxytoca) with urinary tract infection (UTI) in children. We aimed to evaluate the prevalence of fimA, mrkA, matB and pilQ adhesins genes and extended-spectrum beta-lactamase (ESBL) genes blaCTX-M, blaTEM and blaSHV by polymerase chain reaction (PCR) and to study biofilm formation and antibiotics resistance in K. oxytoca from children with UTI.
Methods:
This study was a retrospective cross-sectional study that included 120 children with UTI due to K. oxytoca. The bacteria were subjected to molecular detection of fimA, mrkA, matB and pilQ adhesins genes and ESBL genes blaCTX-M, blaTEM and blaSHV by PCR. Biofilm capacity was determined by the microtiter plate method.
Results:
The isolated K. oxytoca had positive ESBL activity in 45.8% of isolates. About 40% of isolates were biofilm producers. The frequency of adhesion genes among K. oxytoca was 91.7%, 83.3%, 48.3% and 37.5% for matB, pilQ, fimA and mrkA genes, respectively. For ESBL genes, the frequency was 38.3%, 36.7% and 33.3% for blaCTX-M, blaSHV and blaTEM genes, respectively. The commonest genes among ESBL isolates were blaCTX-M (83.6%), blaSHV (80%) then blaTEM gene (72.7%). A significant association (p=0.048) was detected between ESBL activity and biofilm formation by K. oxytoca.
Conclusion:
Present study highlights the emergence of K. oxytoca as a pathogen associated with UTI in children. There was a high prevalence of adhesin genes and ESBL genes among these isolates. The capacity of K. oxytoca to form biofilm was associated with ESBL production.
1. INTRODUCTION
Urinary Tract Infection (UTI) is a global health problem affecting children, common among neonates and infants [1, 2]. The distribution of infection varies with the sex and age of patients. The infection is more common in male neonates and female children, with a global estimated incidence of UTI among girls 3–5%, while among boys, it is 1% [3]. The risk of UTI is due to it is a recurrent infection that can lead to renal scarring after infection in 15-60% of the affected children and renal failure associated with improper management of pyelonephritis [4, 5]
The proper management of UTI includes early diagnosis and proper antibiotics therapy [6]. The proper diagnosis depends upon microbiological urine culture and antibiotic sensitivity tests of the midstream urine sample in older children and clean catch urine sample in infants and younger children [7, 8]. The misuse of antibiotic therapy without available laboratory data is the leading cause of the emergence of extended-spectrum beta-lactamases (ESBL) [9] and carbapenem-resistant Enterobacteriaceae (CRE) [10].
Among Enterobacteriaceae species, Klebsiella oxytoca (K. oxytoca) has emerged as an important pathogen associated with both communities acquired and hospital-acquired UTI [11]. K. oxytoca is a Gram-negative non-motile bacillus that represents a normal intestinal flora in 2%–10% of the population [12, 13]. This species can acquire antibiotic resistance associated with the production of lactamases and carbapenemase enzymes [14].
There are various genes in K. oxytoca that encode ESBL, such as blaCTX-M, blaTEM and blaSHV [15]. Moreover, there are reports about the emergence of carbapenem-resistant K. oxytoca [16]. The emergence of carbapenem resistance among ESBL-producing strains has limited the choice of antibiotics that can be used to treat infection by this organism [17].
Klebsiella oxytoca can produce a biofilm and adhere to the host cells leading to persistence of the infection and antibiotic resistance. There are several adhesins genes associated with the biofilm formation and antibiotic resistance in Klebsiella species [18].
The aims of the present study were to evaluate the prevalence of the fimA, mrkA, matB and pilQ encoding adhesins genes and ESBL genes blaCTX-M, blaTEM and blaSHV by polymerase chain reaction (PCR) and to study the biofilm formation and antibiotics resistance in K. oxytoca isolated from children with urinary tract infections.
2. MATERIALS AND METHODS
2.1. Patients
This study was a retrospective cross-sectional study that included 120 children with a clinical diagnosis of urinary tract infection identified by the urine microbiological culture to be caused by K. oxytoca. The children were recruited from the outpatient clinics of Mansoura University Children Hospital, Egypt, from January 2018 till January 2021.
The inclusion criteria were age below 12 years with one or more symptoms and signs of UTI, including fever above 38°C, urgency, frequency of urine, dysuria and abdominal pain. Children with a previous antibiotic therapy within the last three days or recovered bacterial pathogens other than K. oxytoca were excluded from the study.
The study was approved by the Mansoura Faculty of Medicine Ethical Committee (R.21.03.1272) and confirmed consent was obtained from the parents of each child.
2.2. Bacterial Isolates
2.2.1. Identification of K. oxytoca
The isolates were recovered on MacConkey agar media (Oxoid, Germany) from the urine culture and identified with Gram stain and biochemical reactions by the use of the VITEK 2 system (BioMérieux, Marcy-l'Étoile, France). Further identification was performed by PCR via the amplification of the polygalacturonase (pehX) gene [19].
2.2.2. Antibiotic Sensitivity Testing
The isolated K. oxytoca was subjected to antibiotic sensitivity testing by the disc diffusion method. Briefly, an inoculum from K. oxytoca pure colonies was prepared, adjusted to a 0.5 McFarland standard (1.5 X 108 CFU/ml) and spread over Mueller-Hinton agar plate. The used antibiotic discs were cephalexin (30μg), ceftazidime (30µg), cefepime (30µg), cefotaxime (5μg), cefoxitin (10μg), imipenem (10µg), meropenem (10µg), gentamicin (10µg), tetracycline (10μg), trimethoprim/sulfamethoxazole (25μg) and piperacillin/ tazobactam (36μg) (Oxoid-Thermo Fisher, Hampshire, UK). The antibiotic testing was performed and interpreted according to the Clinical and Laboratory Standards Institute guidelines (CLSI) [20].
2.2.3. Phenotypic Detection of ESBL Enzymes
The isolates were tested for ESBL production with the combined disc test (CDT) according to CLSI guidelines [20]. The combined disc test was performed on ceftazidime- and cefotaxime-resistant strains by placing ceftazidime (30µg) and cefotaxime discs (30µg) at a 20 mm distance from a disc containing ceftazidime/clavulanic acid (CLA) (30/10µg) and cefotaxime/CLA (30/10µg), respectively. If the difference in the inhibition zone diameter was more than 5 mm, these strains were considered ESBL producers.
2.2.4. Phenotypic Detection of Carabapenemase
Carbapenemases production by K. oxytoca was detected by the application of imipenem and meropenem discs with and without 20μl of 0.5M EDTA over Muller-Hinton agar with K. oxytoca isolates and incubated at 37°C for 24 hours. The result was reported positive if the inhibition zone diameter of meropenem was ≤19 mm and the difference in the inhibition zone between meropenem discs with or without EDTA was ≥5 mm [21].
2.2.5. Biofilm Formation by the Microtiter Plate Assay
The ability of K. oxytoca to form a biofilm was studied by the microtiter plate method. The pure colonies of K. oxytoca were subcultured in tryptic soya broth at 37°C for 24 hours. Diluted suspension of the broth was performed in sterile saline at 1:100, then 20 μl was used to inoculate wells of a 96-well flat-bottom polystyrene plate with 180 μl of uninoculated soya broth followed by plate incubation at 37°C for 24 hours. Then, the wells were washed, stained with 10% crystal violet for 15 min, fixed with methanol 99% and left to air-dried for 24 hours. After solubilising with ethanol 96%, the stained wells' optical density (OD) was assessed at 490 nm. The interpretation of the OD was compared with the ODc, which was defined as three standard deviations above the mean OD of the negative control, where OD>4×ODc means strong biofilm formation, OD> 2×ODc and ≤OD4×ODc means moderate biofilm formation, OD ≤2×ODc and >ODc means weak biofilm formation and OD equals the negative control OD means no biofilm formation [22].
2.2.6. Molecular Study of K. oxytoca
2.2.6.1. DNA Extraction
A pure suspension of K. oxytoca colonies was used for DNA extraction by QIAamp DNA Micro Kit (Qiagen, Hilden, Germany).
2.2.6.2. PCR for Detection of ESBL Genes
The blaCTX-M, blaTEM, and blaSHV genes were detected by the PCR assay with specific primers, and 2 µl extracted DNA (Table 1). Klebsiella pneumoniae ATCC 7881 and K. pneumoniae ATCC 700603 strains were used as quality controls. The amplification procedures included denaturation at 95°C for 5 min, followed by 30 cycles of denaturation at 95°C for 30 s, primer annealing at 55°C for both blaTEM and blaSHV or 51°C for blaCTX-M for 30s and primer extension at 72°C for 1 min with final extension 10 min at 72°C [23]. The amplified products were electrophoresed at 1.5% agarose gel for 30 min, and the bands were visualized by ethidium bromide stain using an ultraviolet transilluminator with a DNA marker.
2.2.6.3. PCR for Detection of Adhesin Genes fimA, mrkA, matB, pilQ
The polymerase chain reaction was performed using the primers listed in Table 1, and the amplification procedures included 94°C for 4 min, 30 cycles of 94°C for 30 s, different annealing temperatures for each gene as shown in Table 1 for 30 s, 72°C for 30 s and a final extension of 72°C for 10 min [24]. The amplified products were visualized using 1.5% agarose gel electrophoresis for 30 min and an ultraviolet transilluminator with a DNA marker.
2.3. Statistical Analysis
The data were analyzed by the use of SPSS 22 (Statistical Package of Social Sciences, SPSS Inc, Chicago, Illinois, USA). Numerical data were expressed as median, minimum and maximum as they were non-parametric. The qualitative data were expressed as numbers and percentages. The comparison of qualitative data was performed by Pearson Chi-square test, and the P-value was considered significant if <0.05.
3. RESULTS
This study included 120 children with urinary tract infections recruited from outpatient clinics of Mansoura University Children Hospital from January 2018 - 2021. There were 55% females and 45% males with a minimum age of 0.5 years to 12 years. Their residence was mainly rural (53.3%). The major presenting symptoms were fever and dysuria (46.7% for each), followed by abdominal pain (31.7%) (Table 2).
| Gene | Primer Sequence | Bp | Annealing Temperature |
|---|---|---|---|
| (5/ - 3/) | |||
| blaSHV | F: ATTTGTCGCTTCTTTACTCGC | 1051 | 55°C |
| R: TTTATGGCGTTACCTTTGACC | |||
| blaTEM | F: ATGAGTATTCAACATTTCCGTG | 840 | 55°C |
| R: TTACCAATGCTTAATCAGTGAG | |||
| blaCTX-M | F: TTTGCGATGTGCAGTACCAGTAA | 544 | 51°C |
| R: CGATATCGTTGGTGGTGCCATA | |||
| mrkA | F: CTGGCCGGCGCTACTGCTAAG | 127 | 60 |
| R: CACCCGGGATGATTTTGTTGG | |||
| fimA | F: GCACCGCGATTGACAGC | 132 | 59 |
| R: CGAAGGTTGCGCCATCCAG | |||
| matB/ecp | F: GTACTGGGCGGCAACCTTAG | 98 | 61 |
| R: GTGCCGCTGATGATGGAGAA | |||
| pilQ | F: TCCGCCAGGCTCCACTTC | 194 | 61 |
| R: GCTCGCGGGCATCTGAC |
| Parameter | No. % |
|---|---|
| Age | |
| Mean (± SD) | 5.69 (± 3.96) |
| Minimum | 0.5 |
| Maximum | 12 |
| Median | 5 |
| Percentile | |
| 25% | 2.5 |
| 50% | 5 |
| 75% | 9.87 |
| Sex | |
| Male | 54 45% |
| Female | 66 55% |
| Residence | |
| Urban | 56 46.7% |
| Rural | 64 53.3% |
| Abdominal pain | 38 31.7% |
| Dysuria | 56 46.7% |
| Urgency | 15 12.5% |
| Frequency | 26 21.7% |
| Fever | 56 46.7% |
| Antibiotics | No. % |
|---|---|
| Piperacillin/tazobactam | 48 40% |
| Tetracycline | 45 37.5% |
| Gentamicin | 43 35.8% |
| Trimethoprim/sulfamethoxazole | 58 48.3% |
| Imipenem | 46 38.3% |
| Meropenem | 45 37.5% |
| Cefotaxime | 60 50% |
| Ceftazidime | 65 54.2% |
| Cefepime | 47 39.2% |
| Cephalexin | 70 58.3% |
| Cefoxitin | 57 47.5% |
| ESBL | 55 45.8% |
| Carbapenemase | 43 35.8% |
| Gene | No. % |
|---|---|
| fimA | 58 48.3% |
| mrkA | 45 37.5% |
| matB | 110 91.7% |
| pilQ | 100 83.3% |
| blaCTX-M | 46 38.3% |
| blaSHV | 44 36.7% |
| blaTEM | 40 33.3% |
| Genes |
ESBL Positive Isolates (n=55) No. % |
P-value |
|---|---|---|
| blaCTX-M | 46 83.6% | 0.0001 |
| blaTEM | 40 72.7% | 0.0001 |
| blaSHV | 44 80% | 0.0001 |
| - |
High Biofilm Forming K. oxytoca Isolates (n=3) No. % |
Moderate Biofilm Forming K. oxytoca Isolates (n=45) No. % |
Negative Biofilm Forming K. oxytoca Isolates (n=72) No. % |
P Value* |
|---|---|---|---|---|
| ESBL | 3 100% | 23 51.1% | 29 40.3% | P=0.048 |
| Carbapenemase | 3 100% | 31 68.9% | 44 61.1% | P=0.30 |
| fimA | 2 66.7% | 23 51.1% | 33 45.8% | P=0.697 |
| mrkA | 0 0% | 18 40% | 27 37.5% | P=0.383 |
| matB | 3 100% | 38 84.4% | 69 95.8% | P=0.083 |
| pilQ | 2 66.7% | 34 75.6% | 64 88.9% | P=0.125 |
The isolated K. oxytoca had a high resistance to cephalexin (58.3%), followed by ceftazidime, cefotaxime, Trimethoprim/sulfamethoxazole and cefoxitin; 54.2%, 50%, 48.3% and 47.5%, respectively. The lower resistance of K. oxytoca was for gentamicin (35.8%) and tetracycline (37.5%). The ESBL testing by the combined disc test was positive in 45.8% of the isolated K. oxytoca, and carbapenemase activity was detected by EDTA disc in 35.8% of K. oxytoca isolates (Table 3).
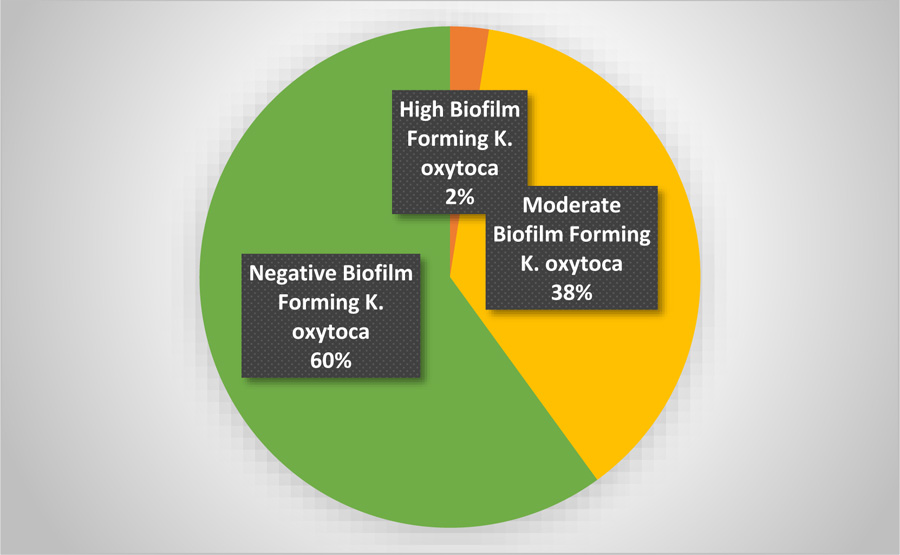
The study of biofilm formation by the microtiter plate assay revealed that 60% of the isolated K. oxytoca was negative, and 40% was positive for biofilm production. The OD of biofilm-forming K. oxytoca revealed that 37.5% had a moderate capacity to form the biofilm, and 2.5% had a high capacity to form the biofilm (Fig. 1).
The frequency of the studied adhesin genes by PCR among the isolated K. oxytoca was 91.7%, 83.3%, 48.3% and 37.5% for matB, pilQ, fimA and mrkA genes, respectively. For ESBL genes, the frequency of their isolation among K. oxytoca isolates was 38.3%, 36.7% and 33.3% for blaCTX-M, blaSHV and blaTEM genes, respectively. The commonest gene among ESBL producing isolates was blaCTX-M (83.6%), followed by blaSHV (80%) and blaTEM gene (72.7%). These ESBL genes were significantly associated with the ESBL K. oxytoca resistance phenotype (P = 0.0001) (Tables 4 and 5).
There was a high frequency of the fimA, mrkA, matB and pilQ adhesin genes in both the moderate and high biofilm-forming K. oxytoca isolates compared to the negative biofilm-forming K. oxytoca isolates. However, this increase was statistically insignificant; P=0.697, 0.383, 0.083 and 0.125 for fimA, mrkA, matB and pilQ genes, respectively. A statistically significant association was detected between ESBL activity and the high and moderate biofilm formation capacities (100% and 51.1%, respectively) among K. oxytoca isolates compared to isolates with no capacity to form a biofilm (40.3%), P=0.048. For carbapenemase activity, it was more prevalent among isolates of K. oxytoca with a high and moderate capacity to form a biofilm (100% and 68.9%, respectively) compared to the isolates with no capacity to form a biofilm (61.1%). However, this association was statistically insignificant (P=0.3) (Table 6).
4. DISCUSSION
Urinary tract infections in children due to bacterial etiology are common and associated with high morbidity. These infections are a common cause of antibiotics prescription, especially for pediatric outpatients [25]. However, the bacterial pathogens associated with these infections have been identified to have a high resistance to the commonly prescribed antibiotics such as cephalosporines [26].
In the present study, UTI was equally common in boys and girls during the first year of life and became more common in girls after the first year of life [27]. The common presenting symptoms were fever, dysuria and abdominal pain. Systemic manifestations such as fever are common with ascending bacterial infection with bacteremia. In children, there is a challenge in clinical diagnosis due to the vague clinical presenting symptoms [28]. Therefore, accurate diagnosis relies upon the laboratory diagnosis with the determination of antibiotics susceptibility pattern [29].
Klebsiella species are an important etiology of infections in children, commonly K. pneumoniae, followed by Klebsiella oxytoca, Klebsiella ozaenae, and Klebsiella rhinoscleromatis [30]. Klebsiella species had emerged as a resistant bacterial pathogen. In the present study, the isolated K. oxytoca had a high resistance to cephalosporines antibiotics. Similar results of resistance to cephalosporines among Klebsiella species and E. coli isolated from urinary tract infections in children were reported previously [30, 31] which may be attributed to the production of various β-lactamase enzymes [32].
On the other hand, in the current study, a lower resistance was reported to gentamicin (35.8%) and tetracycline (37.5%). Studies on the sensitivity of Klebsiella species are contradictory; high resistance to gentamicin was reported in one study [33], while another study revealed high sensitivity to gentamicin and tetracycline [34]. These variations in the sensitivity to different antibiotics may explain the difference in the antibiotics prescribing policy in different geographic locations.
The phenotypic detection of ESBL by the combined disc test revealed that 45.8% of the isolated K. oxytoca was positive. Klebsiella species defined as ESBL producers represent a serious threat to the treatment regimen as this leads to the use of more costly antimicrobials therapy [30]. Production of ESBL is more common in K. pneumonia and appears to be a major factor contributing to drug-resistant UTI among children. These findings call for the need to incorporate ESBL testing in routine clinical practice. The resistance level to commonly prescribed first-line antibiotics observed within Enterobacteriaceae is alarming, necessitating strengthened antimicrobial stewardship [35].
As regard carbapenemase activity of the isolated K. oxytoca, it was positive in 35.8% of the isolates. The treatment of infection caused by bacterial species producing ESBL relies mainly upon the use of carbapenems, including imipenem, meropenem, and ertapenem. However, with the increase of carbapenemase-producing bacterial species, there will be a limitation in the treatment option for infections such as febrile urinary tract infections [36]. The resistance to carbapenems may be attributed to the presence of metallo-β-lactamase and serine carbapenemase enzymes or the presence of other resistance mechanisms such as the loss of outer membrane porins that influence the antibiotic uptake [37]. These findings require further studies about the mechanism of carbapenem resistance among Klebsiella oxytoca isolates.
For ESBL genes, the frequency of blaCTX-M, blaSHV and blaTEM was 38.3%, 36.7% and 33.3%, respectively, and they were highly associated with the ESBL resistance phenotype. The lower prevalence of these genes in different Klebsiella species has been reported in Ghana [38]. The difference can be attributed to the difference in the studied clinical samples and the different studied Klebsiella species.
The presence of adhesive structures enables the bacteria to colonize the host and produce a biofilm and the restriction of antibiotic penetration into the cells. The biofilm formation has been less studied among K. oxytoca isolates. In the current work, the biofilm formation among the isolated K. oxytoca was relatively high; 40% of the isolates had a tendency to form a biofilm either with moderate OD (37.5%) or high OD (2.5%). The ability of isolated K. oxytoca to form a biofilm is relatively lower than that previously reported by Ghasemian et al. for K oxytoca [24]. This might be again attributed to the difference in the source of isolated K. oxytoca from clinical samples and the studied sample size.
MatB, pilQ, fimA, and mrkA were found to be present in 91.7 percent, 83.3 percent, 48.3 percent, and 37.5 percent of the isolated K. oxytoca, respectively, in accordance with another study from Iran [24]. These genes had a higher prevalence among the biofilm-forming K. oxytoca isolates. However, this increase was not statistically significant. Our results support the association between the adhesin genes and biofilm formation. However, complementary studies are needed in this regard due to the lack of previous data related to K. oxytoca strains associated with urinary tract infections in children.
In the present study, there was a significant increase in the ESBL resistance phenotype among the biofilm-forming K. oxytoca isolates. Similar findings have been reported regarding the association between Gram-negative bacilli isolates from urinary tract infections and ESBL [39]. The biofilm formation is associated with changes in the genes expression and metabolic activity of the bacteria leading to the development of antibiotic resistance [40]. The biofilm-producing bacteria might be selected under antibiotic pressure, or the bacteria in the biofilm might acquire antibiotic resistance from exposure to sub-inhibitory antibiotics concentrations; the exact mechanism is still not clear [22]. The biofilm formation and β-lactamases production synergistically contribute to the extensive dissemination of multi-drug resistant strains of Gram-negative bacilli. They are responsible for chronicity, persistence and relapse of infections leading to high morbidity and mortality thus, posing serious health crisis [40].
The favorable therapeutic option for the biofilm-forming isolates will be the use of biofilm remediation therapies. Many of these drugs and strategies offer a promising approach for the future of biofilm therapeutics [41].
CONCLUSION
The present study highlights the emergence of Klebsiella oxytoca as a pathogen associated with urinary tract infections in children. There was a high prevalence of adhesin genes and ESBL genes among these isolates. The capacity of K. oxytoca to form a biofilm was associated with ESBL production. This finding warrants the careful identification of ESBL patterns among the isolates from urinary tract infection in children for an accurate antibiotics prescription policy.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
The study was approved by the ethics committee of Mansoura Faculty of Medicine, Egypt (R.21.03.1272).
HUMAN AND ANIMAL RIGHTS
No animals were used in this research. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or research committee and with the 1975 Declaration of Helsinki, as revised in 2013.
CONSENT FOR PUBLICATION
Informed consent was obtained from the parents of each child.
STANDARDS OF REPORTING
STROBE guidelines and methodologies were followed in this study.
AVAILABILITY OF DATA AND MATERIALS
Not applicable.
FUNDING
None.
CONFLICT OF INTEREST
The authors have no funding or conflicts of interest to disclose.
ACKNOWLEDGEMENTS
Declared none.

