All published articles of this journal are available on ScienceDirect.

The Role of IL-6, TNF-α, and VDR in Inhibiting the Growth of Salmonella Typhi: in vivo Study
Abstract
Background and aim:
The prevalence of typhoid fever is reportedly high, especially in Asia. When a pathogen enters the human body, there are markers in the form of molecules that will be known by the innate immune system. Specific molecular markers of gram negative bacteria, which are Lipopolysaccharides (LPS) and Toll-Like receptors-4 will interact with LPS. The binding between LPS and TLR-4 will give rise to activation signals that will activate innate immune cells. Immune cells will release a number of proinflammatory cytokines, such as TNF-α, IL-1, and IL-6. While Vitamin D Receptors (VDR) are expressed in large amounts in tumor tissue and infected cells. This study aimed to prove the role of IL-6, TNF-α, and VDR in inhibiting bacterial growth in mice that have been induced by S.Typhi.
Methods:
This research was a real experimental pre-post test design to investigate the level of IL-6, TNF-α and VDR in suppressing the growth of bacteria in the peritoneal fluid of S. Typhi, male, mice BALB/c. Mice were divided into three groups comprised of 10 mice each. All mice in groups A and B were intraperitoneally inoculated with S. Typhi strain Thy1 in study day 0. Group A was treated with antibiotic Levofloxacine, on study day 4th. Another study group, group B, was used as a placebo and received aquades on study day 4th. While group C as a control was not inoculated with S. Typhi. Blood samples from three groups for the calculation of serum Il-6, TNF-α, and VDR were collected. This examination was taken four times; at baseline, 4th day, 10th day, and 30th day. For the calculation of bacterial colony, peritoneal fluid retrieval was collected three times, which is on 4th day, 10th day, and 30th day.
Results:
A repeated measure ANOVA in group A (antibiotic) and group B (placebo) group showed that mean IL-6, TNF-α, and VDR level differed statistically significant between times (p-value 0.000). There was a strong negative correlation between bacterial colony count and VDR level, which was statistically significant in both groups (group A; r = -0.875, p-value = 0.000 vs group B; r = -0.470, p-value = 0.002). IL-6 and TNF-α didn't give significant statistical correlation with bacterial colony count.
Conclusion:
VDR, IL-6, and TNF-α play an important role in killing bacteria. From the results of this study, IL-6 level is related to the number of bacterial colonies, the lower the IL-6 level, the less the number of bacterial colonies. Similarly, TNF-α levels have a positive correlation with the number of bacterial colonies. While VDR levels are also related to the number of bacterial colonies, the higher the VDR level, the lower the number of bacterial colonies.
1. INTRODUCTION
Typhoid fever is a systemic infection due to Salmonella Typhi (S.Typhi). In 2011, S.Typhi was estimated to infect 21.7 million people and caused 217,000 deaths worldwide. High incidence of typhoid fever is found in South Asia, Southeast Asia, and South Africa, as many as 80% of cases come from slums in Bangladesh, China, India, Indonesia, Laos, Nepal, Pakistan, and Vietnam [1].
In Indonesia, the prevalence of typhoid fever based on the diagnosis by health professionals and respondent complaints is 1.60%. A total of 14 provinces have Typhoid prevalence above the national prevalence, namely Nanggroe Aceh Darussalam, Bengkulu, West Java, Central Java, Banten, West Nusa Tenggara, East Nusa Tenggara, South Kalimantan, East Kalimantan, Central Sulawesi, South Sulawesi, Gorontalo, West Papua and Papua [2].
Salmonella Typhi is a gram-negative bacillus that causes typhoid fever in humans. This bacterium can survive in the phagosome so that it can escape the body's immune system. Some complications of typhoid fever are ileal perforation, bacteremia, and endovascular infection [3].
When a pathogen enters the human body, there are markers in the form of molecules that will be known by the human defense (innate immune system). Specific molecular markers of gram negative bacteria are lipopolysaccharides (LPS) and Toll-Like Receptors 4 (TLR4) will interact with bacteria (LPS) [4]. The binding between LPS and TLR-4 will give rise to activation signals that will activate innate immune cells. Immune cells will release a number of proinflammatory cytokines, such as TNF-α, IL-1 and IL-6.
IL-6 functions in non-specific and specific immunity produced by mononuclear phagocytes, endothelial cells, vascular, fibroblasts, and other cells in response to other microbes and cytokines. IL-6 is an important member of the cytokine family. It can induce differentiation of T and B lymphocytes. It can also promote the synthesis of acute-phase response proteins by stimulating liver cells. In numerous cytokines, some are very similar in terms of their biological activity [5]. Another study has found that IL-6 levels increase significantly in the early stage of inflammation, which provides evidence for rapid diagnosis and differential diagnosis of early bacterial infection in the clinic [6].
TNF-α plays a role in host defense against bacterial, viral, and parasitic infections. TNF-α is produced by macrophages and is activated by lymphocyte T cells, antigens, NK cells, and mast cells [7]. TNF-α is usually not detected in healthy individuals but is often found in conditions of inflammation and infection in the serum. TNF-α acts on leukocytes and endothelial, inducing acute inflammation at low levels because TNF-α is a strong pyrogen [8, 9].
Vitamin D receptors or VDR are expressed in large amounts in tumor tissue and infected cells. Recent research shows that VDR and enzymes involved in vitamin D metabolism have damage to the VDR signaling pathway. VDR expression by immune cells shows that vitamin D influences the immune system function. More than 30 different body tissues, such as the brain, liver, and pancreas, lymphatics, skin, gonads and prostate are composed of cells including T and B lymphocytes that express VDR. Vitamin D The receptor binds to 1,25-hydroxyvitamin D, the active form of vitamin D and mediates its biological activity [10]. Based on various previous theories, this study aimed the role of IL-6, TNF-α, and VDR in inhibiting bacterial growth in mice that have been induced by S.Typhi.
2. MATERIALS AND METHODS
This research is a true experimental pre-post test design to investigate the level of IL-6, TNF-α, and VDR in suppressing the growth of bacteria in the peritoneal fluid of S. Typhi strain male mice BALB/c.
2.1. Experimental Animals
BALB/c mice (male, aged 8-12 weeks, weighing 30-40 grams; n = 30) were maintained in the Molecular Biology and Immunology Laboratory, Microbiology Department Faculty of Medicine, Hasanuddin University (Makassar, Indonesia). The mice were acclimatized for 8 days. Randomization was used as a sampling technique. Mice were randomly put into three study groups of 10 mice each. Two groups (A and B) were intraperitoneally induced with S. Typhi strains thy1 (3 mL x 103 CFU/mL). Group A was treated with antibiotic levofloxacin once a day for five days in a dose of 1.95 mg/kg, and group B received placebo treatment. Control group C did not receive either S. Typhi nor antibiotic.
2.2. Levofloxacin
Levofloxacin was obtained from Kimia Farma, Pharmaceutical, Indonesia. Dose 750 mg of Levofloxacin given to mice was obtained from the multiplication with a conversion factor of 0.0026. Based on the result, the positive control group was given a dose of Levofloxacin 1.95 mg/day. Antibiotics was dissolved in distilled water and given through the nasogastric sonde, once a day for five days.
2.3. Sampling of Peritoneal Fluid And Bacterial Colony Examination
Mice were fixed in the supine position, the abdomen is cleaned with alcohol 70% and as much as 0.8-1 mL saline was injected into the peritoneal cavity. Mice were then allowed to stand for 1 minute as he rocked slowly. Peritoneal fluid removed from the peritoneal cavity of mice supine position, then fluid aspirated by as much as 0.5 mL syringe. Peritoneal fluid retrieval is performed three times, which is on the fourth day (4th day), on the tenth day (10th day) and 3 weeks after the mice induced with S. Typhi (30th day).
Examination of bacterial colonization by using the pour plate method. This method is performed by diluting the peritoneal fluid samples of 0.5 mL in 4.5 mL of saline (0.9% NaCl). Dilution is done three times so that the culture obtained is not too dense or fulfill cup (culture too dense will interfere with observations). Approximately, 1 mL of the suspension was poured into a sterile petri dish, followed by pouring the fertilizer medium (nutrient agar) sterile warm (450C), then sealed and incubated for 1-2 days at a temperature of 370C.
2.4. Il-6, TNF-α And VDR Examination
Blood samples from two groups for the determination of serum Il-6, TNF-α, and VDR were collected. This examination was taken four times; at baseline, 4th day, 10th day and 30th day. The examination was done using IL-6 ELISA kit (Sandwich ELISA), TNF-α ELISA Kit (Sandwich ELISA), VDR ELISA Kit (Sandwich ELISA) for mouse, purchased from LSBio.
2.5. Statistical Analysis
The data obtained is processed using SPSS 23 (IBM Corporation, New York, USA) for Macbook. All data are presented in the form of a mean and standard deviation. Before testing the difference hypothesis, first, we test the data normality and homogeneity. One-way ANOVA is used to calculate the mean difference of Il-6, TNF-α and VDR level between antibiotic group, placebo group, and control group at baseline. Because in this study, each group is measured multiple times to see changes to specific intervention, so we also used ANOVA with a repeated measure to calculate the differences means of bacterial colony count, Il-6, TNF-α and VDR level in the same groups in multiple time observation. ANOVA repeated measure would be followed by Bonferroni post hoc test to identify which particular differences pairs of groups means are significant. Considered as statistical significance was p-value <0.05. A Pearson correlation test was used to determine the correlation between bacterial colony count with VDR, IL-6, and TNF-α. Linear regression analysis was used to determine the most influential variables in determining the bacterial colony count.
3. RESULTS
This study consisted of 30 mice, which were divided into 3 groups (n = 5). Data on the characteristics of mice can be seen in Table 1. No significant differences were found for age, body weight, VDR, Il-6 and TNF-α level at baseline (Table 1).
The result found that at baseline, there was no statistically significant difference between VDR level in three groups. Both IL-6 and TNF-α levels also give no statistically significant differences in mean (Table 1).
A repeated measure ANOVA in group A (antibiotic) and group B (placebo) showed that mean VDR level differed statistically significant between times (p-value 0.000), while in control group C showed no significant differences (p-value 0.495). There are significant differences in the average increase in VDR levels over time in group A (antibiotic) and group B (placebo). Increased levels of VDR were observed to be greater in the group with antibiotics compared to the placebo group and control group. Post hoc tests using Bonferroni correction revealed that in group A (antibiotic) shows an increase in VDR level from 4th day to 10th day (6.95±1.32 ng/mL vs. 15.04±2.37 ng/mL, p-value 0.000), which was statistically significant. In group B (placebo), post hoc tests using Bonferroni correction revealed that in this group shows an insignificant decrease in VDR level from 4th day to 10th day (7.95±1.71 ng/mL vs. 7.93±1.42 ng/mL, p-value 1.000). Therefore, we can conclude that the group with given antibiotics can induce increase of VDR level more significant than with placebo (Table 2).
A repeated measure ANOVA in group A (antibiotic) and group B (placebo) showed that the mean IL-6 level differed statistically significant between times (p-value 0.000), while in control group C showed no significant differences (p-value 0.055). Post hoc tests using Bonferroni correction revealed that in group A (antibiotic) shows an increase in IL-6 level from baseline day to 4th day (109.25±29.21 pg/mL vs 481.72±44.10 pg/mL, p-value 0.000), which was statistically significant. The same result was found in the group B (placebo), it shows an increase in IL-6 level from baseline day to 4th day (125.71±40.12 pg/mL vs 248.87±32.75 pg/mL, p-value 0.000). Given antibiotics for five days, 4th day to 10th day, showed nonsignificant decreasing in IL-6 (481.72±44.10 pg/mL vs 465.58±45.85 pg/mL, p-value 1.000). In group B (placebo), post hoc tests using Bonferroni correction revealed that an insignificant decrease in IL-6 level from 4th day to 10th day (248.87±32.75 pg/mL vs 222.84±37.20 pg/mL, p-value 0.326) is shown in this group. Therefore, we can conclude that a group that induced infection can induce increasing IL-6 level. After the intervention, both groups given antibiotic and placebo showed a decrease in IL-6 (Table 2).
A repeated measure ANOVA in group A (antibiotic) and group B (placebo) showed that the mean TNF-α level differed statistically significant between times (p-value 0.000). In contrast, in control group C showed insignificant differences (p-value 0.801). Post hoc tests using Bonferroni correction revealed that in group A (antibiotic) shows an increase in TNF-α level from baseline day to 4th day (96.62±21.90 pg/mL vs 304.88±38.31 pg/mL, p-value 0.000). The same result was found in the group B (placebo), it shows an increase in TNF-α level from baseline day to 4th day (98.96±25.95 pg/mL vs 198.46±34.65 pg/mL, p-value 0.001). Post hoc tests using Bonferroni correction revealed that in antibiotic group shows a decrease in TNF-α level from 4th day to 10th day (304.88±38.31 pg/mL vs 294.01±24.34 pg/mL, p-value 1.000), which was statistically not significant. In group B (placebo), post hoc tests using Bonferroni correction revealed that in this group also shows insignificant decrease in TNF-α level from 4th day to 10th day (198.46±34.65 pg/mL vs 183.73±10.80 pg/mL, p-value 0.929). Therefore, we can conclude that group that induced with infection can induce increasing TNF-α level. After the intervention, both groups were given antibiotics and placebo showed a decrease in TNF-α (Table 2).
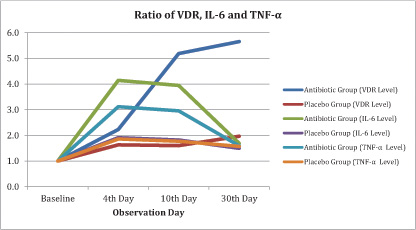
Fig. (1) showed that there was an increase in VDR levels in group A (antibiotic) and group B (placebo), with reasonably wide differences in mean. While IL-6 and TNF-α levels increased on the 4th day and then decreased after the 10th day.
| Variable | Groups | - | p-value | |
|---|---|---|---|---|
| Group A (antibiotic) (n=10) |
Group B (placebo) (n=10) |
Group C (control) (n=10) |
- | |
| Strain | BALB/c | BALB/c | BALB/c | - |
| Sex | male | male | male | - |
| Age (weeks) | 10.00±1.49 | 9.60±1.43 | 9.90±1.28 | 0.804 |
| Body weight (grams) | 34.54±3.45 | 35.72±2.23 | 34.36±3.65 | 0.614 |
| VDR level (ng/mL) at baseline | 2.98±1.09 | 3.82±1.81 | 2.94±1.07 | 0.275 |
| IL-6 level (pg/mL) at baseline | 109.25±29.21 | 125.71±40.12 | 103.55±22.98 | 0.282 |
| TNF-α level (pg/mL) at baseline | 96.62±21.90 | 98.96±25.95 | 95.50±20.01 | 0.909 |
| Groups | VDR level (ng/mL) | ||||
|---|---|---|---|---|---|
| Baseline | 4th day | 10th day | 30th day | p value* | |
| Group A (antibiotic) | 2.98±1.09 | 6.95±1.32 | 15.04±2.37 | 16.35±2.27 | 0.000 |
| Group B (placebo) | 3.82±1.81 | 7.95±1.71 | 7.93±1.42 | 9.28±1.51 | 0.000 |
| Group C (Control) | 2.94±1.07 | 3.09±1.42 | 2.63±1.12 | 3.06±0.96 | 0.495 |
| Groups | IL-6 level (pg/mL) | p value* | |||
| Group A (antibiotic) | 109.25±29.21 | 481.72±44.10 | 465.58±45.85 | 198.90±31.49 | 0.000 |
| Group B (placebo) | 125.71±40.12 | 248.87±32.75 | 222.84±37.20 | 197.34±39.89 | 0.000 |
| Group C (Control) | 103.55±22.98 | 112.25±27.96 | 89.63±19.68 | 85.71±17.27 | 0.055 |
| Groups | TNF-α level (pg/mL) | p value* | |||
| Group A (antibiotic) | 96.62±21.90 | 304.88±38.31 | 294.01±24.34 | 168.35±19.09 | 0.000 |
| Group B (placebo) | 98.96±25.95 | 198.46±34.65 | 183.73±10.80 | 157.77±21.59 | 0.000 |
| Group C (Control) | 95.50±20.01 | 95.60±21.86 | 91.13±16.95 | 93.98±15.26 | 0.801 |
| Groups | Bacterial Colony Count (CFU/mL) | |||
| 4th day | 10th day | 30th day | p value* | |
| Group A (antibiotic) | 20.20±4.96 | 0.70±1.33 | 0.00±0.00 | 0.000 |
| Group B (placebo) | 21.90±5.93 | 6.10±2.07 | 1.10±1.10 | 0.000 |
| Variable | Group A (antibiotic) | Group B (placebo) | ||
| Bacterial colony (CFU/mL) | Bacterial colony (CFU/mL) | |||
| r-value | p value* | r-value | p value* | |
| VDR (ng/mL) | -0.875 | < 0.001 | -0.470 | < 0.05 |
| IL-6 (pg/mL) | -0.069 | > 0.05 | -0.203 | > 0.05 |
| TNF-α (pg/mL) | -0.150 | > 0.05 | -0.180 | > 0.05 |
| Variable | Group A (antibiotic) | Group B (placebo) | ||||
| Standard Error | Β-Coefficients | p value* | Standard Error | Β-Coefficients | p value* | |
| VDR (ng/mL) | 0.145 | -1.623 | 0.000 | 0.752 | -0.557 | 0.005 |
| IL-6 (pg/mL) | 0.012 | 0.016 | 0.190 | 0.037 | 0.066 | 0.757 |
| TNF-α (pg/mL) | 0.023 | -0.014 | 0.557 | 0.047 | 0.086 | 0.681 |
A repeated measure ANOVA in group A (antibiotic) and group B (placebo) showed that mean bacterial colony count differed statistically significant between times (p-value 0.000). Post hoc tests using Bonferroni correction revealed that in group A (antibiotic) shows a decrease in bacterial colony count from 4th day to 10th day (20.20±4.96 CFU/mL vs 0.70±1.33 CFU/mL, p-value 0.000), which was statistically significant. While there is a non-significant decrease in bacterial colony count from 10th day to 30th day (0.70±1.33 CFU/mL vs 0.00±0.00 CFU/mL, p-value 0.397). In group B (placebo), post hoc tests using Bonferroni correction revealed that in this group, decreasing significant decrease in bacterial colony count from 4th day to 10th day (21.90±5.93 CFU/mL vs 6.10±2.07 CFU/mL, p-value 0.000), also between 10th day to 30th day (6.10±2.07 CFU/mL vs 1.10±1.10 CFU/mL, p-value 0.000) is observed. Therefore, we can conclude that both groups can show decrease in bacterial colony count, even in the placebo group. We can assume the role of innate immunity that can induce cytokins, such as IL-6 or TNF-α that help to fight against the bacteria (Table 3).
A Pearson correlation was used to determine the relationship between bacterial colony count and other variables. Based on the result, there was a strong negative correlation between bacterial colony count and VDR level, which was statistically significant in both groups (group A; r = -0.875, p-value = 0.000 vs group B; r = -0.470, p-value = 0.002). IL-6 and TNF-α didn't give a significant correlation with bacterial colony count (Table 4).
Multiple regression was used to predict the correlation between bacterial colony count from VDR, IL-6 and TNF-α level in the group given antibiotic and placebo. The result of multiple linear regression analysis showed that VDR plays a role in determining in a decrease in bacterial colony count, while other variables were not significant. The higher VDR level, the lower bacterial colony count in both group (group A (antibiotic) ; Β-Coefficients = -1.623, p-value = 0.000 vs group B (placebo) ; Β-Coefficients = 0.557, p-value = 0.005) (Table 5).
4. DISCUSSION
When S. Typhi first enters the body, bacteria will be destroyed by macrophages. Bacteria will be known by various receptors located on the surface of phagocytes [11]. Specific marker molecules for gram-negative bacteria, such as S. Typhi are Lipopolisakarida (LPS). LPS will activate TLR-4, which is a receptor that plays a role in observing and destroying S. Typhi. Activated TLR-4 will recruit the MyD88 adapter protein. Then, MyD88 recruited IRAK4, IRAK1 and IRAK2. IRAQ kinase then phosphorylates and activates the TRAF6 protein, allowing NF-κB to dwell in the cell nucleus and activating transcription and causing induced inflammatory cytokines. Proinflammatory cytokines, such as IL-1β and IL-6, IFN-γ and TNF-α are synthesized and systemic inflammation occurs [12].
In this study, we found a significant increase in IL-6 levels on the 4th day compared to baseline because the S. Typhi bacteria had been injected since day 1. In this study, a group of mice in the given antibiotic and placebo, showed an increase in mean levels of IL-6 at 4th day compared to baseline (Table 2). It can be concluded, after induction by S. Typhi, the inflammatory process by cytokines IL-6 has been running. After cytokines are secreted, the helper-1 (Th1) and T helper-2 (Th2) will be activated. Signals from cytokines triggered by the interaction of host cells and bacteria are crucial in the development of the disease. The balance between proinflammatory and anti-inflammatory cytokines will control the prevention of host damage due to excessive inflammation [13].
After giving antibiotics for 5 days on, the average value of IL-6 levels decreased in the antibiotic group (Table 2). Antibiotics are given that can kill bacteria, so it occurs modulating the LPS binding pathway with TLR-4 by the death of bacteria. LPS in bacterial cell walls is reduced so that the TLR-4 protein is not activated. Then by the mechanism of action to modulate the transcription factor pathways, such as NF-κB, so that the production process of IL-6 inflammatory cytokines becomes inhibited and IL-6 levels decrease.
In this study, we calculated the amount of bacterial colonization through the sampling of peritoneal fluid and found a decrease in mean levels of bacterial colony at 10th day compare to 4th day (Table 3). S. Typhi infection given intraperitoneum will stimulate macrophages to activate and move to the source of infection. LPS content in S. Typhi cell walls is a signal for macrophages to activate. Activation of macrophages has a high ability to ingest foreign objects through phagocytosis. These cells will destroy all foreign objects, such as bacteria, damaged cells, tumor cells, colloidal objects, and large molecules [14][15]. The increasing number of macrophages to the site of infection originates from the migration of macrophages to the source of stimulation. In addition, the increase in the number of macrophages is caused by the acceleration of the proliferation and differentiation of macrophages. Proliferation and differentiation, as well as increased migration of macrophages to the source of stimulation cause the number of macrophages in the peritoneum to increase. The acceleration of migration is also due to stimulation by IL-6 produced by monocyte cells or macrophages [15]. The IL-6 level on the 30th day showed that the value decreased even more along with the number of bacterial colonization (Fig. 1).
TNF-α is a pro-inflammatory cytokine that is produced most by macrophages through several mechanisms [16][17]. Immunologically, TNF-α is produced through macrophage presentation to the Antigen-Presenting Cell (APC) which then APC will order T-helper 1 to proliferate into IL-12 and IFN-γ where IFN-γ will produce TNF-α as an antimycobacterial. Other studies also explain the same thing, the process of infection can cause plasma enlargement by endothelial activation, so that infected magrofag will become active and release cytokines including tumor necrosis factor alpha (TNF-α), interleukin 1 (IL-1), IL- 6, and platelet activating factor (PAF) [18]. In this study, group A that given antibiotics showed that the mean TNF-α level differed statistically significant between times (p-value 0.000) (Table 2). This is because the antibiotics given to the mice in group A can reduce the number of S. Typhi thereby suppressing the infection process in the host directly, causing a reduction in cytokine release by magrofag, one of which is TNF-α. Whereas TNF-α levels in the group B that given placebo, although decreased on 10th day and 30th day, the mean decrease was lower than in the group with antibiotics.
Vitamin D receptors are expressed in large amounts in the tumor tissue and the infected cells. Recent research has shown VDR and enzymes involved in the metabolism of vitamin D have damage to the VDR signaling pathways. VDR expression by immune cells suggests that vitamin D affects the immune system function. More than 30 different body tissues, such as the brain, liver, and pancreas, lymphatic, skin, gonads, and prostate consists of cells including T and B lymphocytes that express the VDR. Vitamin D receptor binds to the 1,25(OH)2D3, the active form of Vitamin D and the mediated its biological effects [10]. Dendritic cells are the primary targets for the immunomodulatory activity of 1,25(OH)2D3, by inhibiting the differentiation and maturation of dendritic cells, suppressing the expression regulation of MHC-II, costimulatory molecules (CD40, CD80 and CD86) and decreased production of IL-12. In addition, 1,25(OH)2D3 increases the production of IL-10 and promote apoptosis dendritic cells. Together, the effect of 1,25(OH)2D3 inhibits the activation of T cells that depend on DC [19]. In this study, a group of mice that given antibiotic in group A, showed an increase in mean levels of VDR on 4th day compared to 10th day (Table 2). VDR activation process then activates the transcription factor genes antimicrobial natural peptide, cathelicidin, and defensins [20], then later will kill the bacteria. This was proved in the result that there was a strong negative correlation between bacterial colony count and VDR level (Table 4). Mice that do not have the VDR impaired production of Th1 promotive factor and IL-18, decreased Th1 cell proliferation and decreased expression of signal transducer and activation of transcription 4 (STAT 4) (a transcription factor Th1 cells). Taken together, these circumstances indicate that VDR function is essential for the development of Th1 cells. In mice that did not have VDR, will have a decreased proliferative response to stimulation of CD3 [19][21][22].
CONCLUSION
VDR, IL-6, and TNF-α play an important role in killing bacteria. From the results of this study, IL-6 levels are related to the number of bacterial colonies, the lower the IL-6 level, the less the number of bacterial colonies. Similarly, TNF-α level has a positive correlation with the number of bacterial colonies. While VDR level is also related to the number of bacterial colonies, the higher the VDR level, the lower the number of bacterial colonies.
ABBREVIATION USED
| CFU | = Colony Forming Units |
| IL-6 | = Interleukin 6 |
| TNF-α | = Tumor Necrosis Factor Alpha |
| VDR | = Vitamin D Receptor |
ETHIC APPROVAL AND CONSENT TO PARTICIPATE
This study was approved by the Faculty of Medicine, Hasanuddin University Makassar, Indonesia, with registration number 901 / H4.8.4.5.31 / PP36-KOMETIK / 2018.
HUMAN AND ANIMAL RIGHTS
No humans were used. All experiments on animals were in accordance with the Health Medical Research Ethics Committee at the Faculty of Medicine, Hasanuddin University (Makassar, Indonesia) as the local guidelines for the care and use of laboratory animals.
CONSENT FOR PUBLICATION
Not applicable.
AVAILABILITY OF DATA AND MATERIALS
The authors confirm that the data supporting the findings of this study are available within the article.
FUNDING
This research was financially supported by the Ministry of Research and Technology, Indonesia.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
Authors would like to thank Rommy Usman, Mus, Wani, and Mark (Molecular Biology and Immunology Laboratory for Infection Diseases, Faculty of Medicine, Hasanuddin University, Makassar, Indonesia) who helped in the implementation of our research activities.

