All published articles of this journal are available on ScienceDirect.

DNA Sequence Analysis of BlaVEB Gene Encoding Multi-drug Resistant and Extended-spectrum β-lactamases Producer Isolates of Enterobacteriaceae and Pseudomonas aeruginosa
Abstract
Objective:
Multi-drug resistance Gram-negative bacteria possessing Extended-Spectrum β-Lactamase (ESBL) genes are of concern because of their resistance to third-generation cephalosporins. This study aims to investigate the molecular basis of resistance to modern β-lactams by ESBLs encoded by the blaVEB gene and the gene’s role in resistance. Also, gene sequencing was used to compare genetic similarities with global isolates using phylogenetic and cluster analyses.
Methods:
Between March and July 2018, a total of 100 Iraqi clinical isolates were examined, in this cross-sectional study, to determine their ESBL status using the double-disc synergy technique. Polymerase Chain Reactions (PCRs) were performed on extracted blaVEB genes and sequencing of the target PCR products was performed. All blaVEB sequences were compared with the available sequence data, using BLAST searches against the GenBank database.
Results:
A total of 35 isolates, comprising 5 Escherichia coli, 18 Klebsiella pneumoniae, and 12 Pseudomonas aeruginosa isolates were confirmed to possess ESBLs; the blaVEB gene was detected in one isolate of each species. The sequencing of these genes revealed 99% similarity with the global standard genes deposited in GenBank.
Conclusion:
The blaVEB gene plays an essential role in the resistance of ESBL-producing isolates to new β-lactams. Further, the sequencing and phylogenetic analyses of the genes from the P. aeruginosa, K. pneumonia, and E. coli isolates revealed 99% similarity with the GenBank global standard genes.
1. INTRODUCTION
Clinically, the most important β-lactam resistance mechanism operating in Gram-negative pathogens is enzymatic antibiotic inactivation by β-lactamases [1]. Extended-spectrum β-lactamase (ESBL)-mediated resistance has become prevalent, worldwide, resulting in Gram-negative bacteria being able to utilize these plasmid-encoded enzymes to hydrolyze the extended-spectrum cephalosporins (e.g., ceftazidime and cefotaxime) [2, 3]Evolutionary pressure has resulted in approximately 1000 different types of β-lactamases that are able to hydrolyze the β-lactam ring of susceptible antibiotics [4]. The plasmids that normally carry these resistance genes play important roles in the transfer of resistance between bacteria, increasing the breadth of the resistance to these and other antibiotics, like gentamicin. Thus, the therapeutic choices available for combatting resistant bacteria remain limited [5].
Among ESBLs, the most widespread and clinically relevant classes are the TEM, SHV, and CTX-M types. The TEM and SHV types are derived from the TEM-1, TEM-2, and SHV-1 penicillinases [6]. Other plasmid-mediated ESBLs, such as PER, VEB, GES, and IBC β-lactamases, have been described but are uncommon and have been found mainly in Pseudomonas aeruginosa and at a limited number of geographic sites [7]. The blaVEB group is one of the smaller subsets of the class A β-lactamases. The blaVEB enzymes appear to be frequently observed in non-fermenting species such as P. aeruginosa, Acinetobacter baumannii, and other Enterobacteriaceae spp., and their rate of proliferation is increasing [8]. This study characterized ESBL-producing Gram-negative bacteria, isolated in Iraq, using primary screening tests and confirmatory phenotypic analyses. Further, the blaVEB genes encoding the ESBLs were detected, sequenced, and compared with other global isolates.
2. MATERIALS AND METHODS
Between March and July 2018, Gram-negative isolates were collected from patients attending the Al-Karkh General Hospital (Baghdad, Iraq). In this cross-sectional study, the bacteria were isolated from patients with burn wounds, non-burn wounds, otitis media, and urinary tract infections. Patient information (sex, age, duration of hospitalization, disease characteristics, type of therapy) was recorded. The bacterial isolates were identified using their morphological and biochemical characteristics. This study was conducted according to the principles of the Declaration of Helsinki; the nature of the study did not require having the patients provide informed consent as there were no patient manipulations involved in the study.
2.1. Preservation and Maintenance of Bacterial Isolates.
Isolates of P. aeruginosa, Escherichia coli, and Klebsiella pneumoniae were derived from patient samples. Bacterial isolates were stored, in a brain heart infusion broth containing 20% glycerol,[9] at -20°C for 6–8 months [9]. Prior to use in susceptibility tests, the bacterial isolates were grown and adjusted to a density equivalent to a 0.5 McFarland Standard [10].
2.1.1. Antimicrobial susceptibility testing
The susceptibilities of the collected isolates to ceftriaxone (30 µg), ceftazidime (30 µg), imipenem (10 µg), meropenem (10 µg), cefoxitin (30 µg), aztreonam (10 µg), and cefepime (30 µg) were determined using the disc diffusion method, as recommended by the Clinical and Laboratory Standards Institute (CLSI) [9].
2.2. Detection of ESBLs
2.2.1. Primary ESBL screening technique
The Kirby-Bauer disc diffusion technique was used to conduct primary ESBL screening. Briefly, this method entails the preparation of a suspension of the target bacteria at a standardized density. The bacterial suspension is used to swab the surface of Mueller-Hinton agar plates, after which antibiotic-impregnated discs were aseptically placed on the agar surface and the bacteria were allowed to grow for 18 h. Thereafter, the resultant zones of inhibition were measured and interpreted, according to CLSI guidelines [9].
2.2.2. Confirmatory ESBL test
In this test, one Amoxiclav (amoxicillin (30 µg) + clavulanic acid (10 μg) disc and one ceftazidime disc (30 μg) were placed on target bacteria-seeded agar plates, 10 mm apart. Amoxiclav and cefotaxime (30 μg) discs were also placed on the same agar plate, the same distance apart. The plates were incubated at 37°C for 16-18 h. An isolate was considered to produce an ESBL if the zone of inhibition around the antibiotic test disc was increased nearer the Amoxiclav disc, according to CLSI [9].
2.3. DNA Extraction
An automatic nucleic acid extraction system (SaMag-12, Sacace Biotechnologies, Como, Italy) was used to extract the genomic DNA from study isolates. For each bacterial isolate, colonies were grown on agar plates, suspended in brain heart infusion broth, and processed according to the manufacturer’s instructions.
2.4. Molecular Technique for the Detection of ESBLs
Isolated genomic DNA samples were used as templates for specific Polymerase Chain Reaction (PCR) amplification and detection of the blaVEB gene. Specifically, the blaVEB forward (5'-CGACTTCCATTTCCCGATGC-3’) and reverse (5’-GGACTCTGCAACAAATACGC-3’) primers were used in conjunction with Maxime PCR Pre-Mix kits (Intron Biotechnologies, Seongnam, Korea). The optimum conditions for detection of the blaVEB gene included initial denaturation at 95°C for 3 min followed by 30 cycles each, of denaturation (95°C for 45 s), annealing (50°C for 45 s), and amplification (72°C for 45 s); a final extension step (72°C for 7 min) completed the reaction. Agarose gel electrophoresis was performed to determine DNA amplicon sizes after extraction; standard DNA ladders were used to determine the size of the DNA bands [11]. The electrophoresis proceeded at 5 V/cm2, in TBE buffer, for 1.5 h. When electrophoresis was completed, each gel was read using ultraviolet light (336 nm).
2.5. DNA Sequencing
Sequencing of the target PCR products was performed by the National Instrumentation Center for Environmental Management (Seoul, Korea). All the blaVEB sequences were compared with the available sequence data, using BLAST searches, against the GenBank database (National Center for Biotechnology Information, Bethesda, MD, USA) to identify the sequences. Multiple sequence alignment was manually performed with closely related reference sequences from other E. coli, K. pneumonia, and P. aeruginosa isolates, available in GenBank, using the BLAST (http://www.ncbi.nlm.nih.gov) and BioEdit (Ibis Therapeutics, Carlsbad, CA, USA) programs. A neighbor-joining tree with combined blaVEB data was constructed, according to maximum likelihood method, using Molecular Evolutionary Genetics Analysis (MEGA) 7 software (ver. 7, Pennsylvania State University, State College, PA, USA) [12].
2.6. Statistical Analysis
The data were analyzed using Excel (Microsoft, Redmond, WA, USA), Minitab (ver. 17, Minitab, State College, PA, USA), and SPSS (ver. 24, IBM, Armonk, NY, USA). The Chi-square test was used to investigate the association between antibiotic susceptibility (sensitivity, resistance, intermediate), ESBL production, and blaVEB gene prevalence among the bacterial isolates; P-values < 0.05 were considered statistically significant.
3. RESULTS
A total of 100 clinical specimens were collected during this study. Among these specimens, 65 were culture-positive for Gram-negative bacteria, 12 for Gram-positive bacteria, and 23 were culture negative. Gram-negative bacteria were isolated from the sites listed in Table 1.
These isolates, screened using the Kirby-Bauer disc diffusion method, showed 21 (72.4%) P. aeruginosa isolates were resistant to ceftriaxone, 19 (65.5%) to ceftazidime, 7 (24.1%) to aztreonam, 13 (44.8%) to cefepime, 29 (100%) to cefoxitin and 1 (3.4%) to imipenem. The K. pneumoniae isolates were resistant to ceftriaxone 24 (92.3%), ceftazidime 23 (88.4%), aztreonam 21 (80.7%), and cefepime 12 (46.2%). The E. coli isolates demonstrated resistance to ceftriaxone 10(100%), ceftazidime 9 (90%), aztreonam 10 (100%), cefoxitin 10 (100%), meropenem 2 (20%), and imipenem 3 (30%).
Based on the confirmatory testing for the extended-spectrum β-lactamase production, 35 isolates were considered to produce ESBLs, including 12 (34%) P. aeruginosa, 18 (51%) K. pneumoniae, and 5 (14%) E. coli isolates.
PCR was then used to detect the presence of the blaVEB gene in the K. pneumoniae, E. coli, and P. aeruginosa clinical isolates. Three isolates, one from each of the three species, were positive for the presence of blaVEB.
3.1. Sequencing and Phylogenetic Analysis
Among the isolates, 3 possessed the blaVEB gene and were sent for sequencing and phylogenetic analysis (Fig. 1). The alignment study of the blaVEB gene in K. pneumoniae revealed a genotype similar to other strains, from Australia, China, Brazil, Greece, and Tunisia, deposited in the GenBank (accession numbers documented in Table 2 and Fig. (2). Sequencing of the gene demonstrated 99% compatibility with the global standard gene in Gen Bank.
| Bacteria | Burn Woundsn (%) | Non-Burn Woundsn (%) | Urinary Tractn (%) | Otitis Media (%) | No. of Isolates n (%) |
|---|---|---|---|---|---|
| K. pneumoniae | 20 (77%) | – | 5 (19%) | 1 (4%) | 26 (40%) |
| P. aeruginosa | 18 (62%) | 2 (7%) | – | 9 (31%) | 29 (45%) |
| E. coli | 4 (40%) | – | 6 (60%) | – | 10 (15%) |
| – | – | – | – | – | 65 |
| Isolate | Accession | Country | Similarity |
|---|---|---|---|
| K. pneumoniae | ID: CP031802.1 | Australia | 99% |
| – | ID: MF417536.1 | Brazil | 99% |
| – | ID: KY630495.1 | Brazil | 99% |
| – | ID: CP006657.1 | China | 99% |
| – | ID: JN406319.1 | Greece | 99% |
| – | ID: KP007363.1 | Tunisia | 99% |
| P. aeruginosa | ID: KX857137.1 | Kenya | 99% |
| – | ID: NG_050326.1 | USA | 99% |
| – | ID: KT283238.1 | India | 99% |
| – | ID: KM094182.1 | Iran | 99% |
| – | ID: HM246150.1 | Singapore | 99% |
| – | ID: HM240861.1 | Iran: Kerman | 99% |
| – | ID: GQ388247.1 | France | 99% |
| – | ID: DQ333895.1 | China | 99% |
| – | ID: MG188749.1 | Brazil | 99% |
| E. coli | ID: NG055500.1 | Thailand | 99% |
| – | ID: NG_050317.1 | USA | 99% |
| – | ID: AF205943.1 | France | 99% |
| – | ID: AF010416.1 | France | 99% |
| – | ID: NG_050323.1 | Norway | 99% |
| – | ID: NG_050325.1 | Thailand | 99% |

| Accession No. | Nucleotide | Identity | Location | Substitution Type | Source Organism |
|---|---|---|---|---|---|
| KM094182.1 | C>G | 99% | 281 | Transversion | P. aeruginos |
| C>T | 294 | Transition | |||
| A>G | 456 | Transition | |||
| A>G | 536 | Transition | |||
| G>C | 547 | Transversion | |||
| MF417536.1 | C>G | 99% | 170 | Transversion | K. pneumonia |
| A>G | 304 | Transition | |||
| C>G | 343 | Transversion | |||
| NG-055500.1 | T>G | 99% | 299 | Transversion | E. coli |
| G>C | 392 | Transversion |
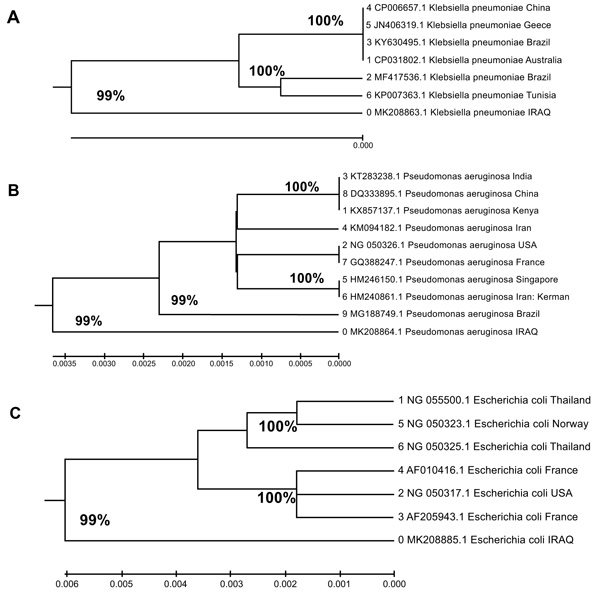
The blaVEB gene that encoded the P. aeruginosa ESBL also demonstrated a genotype closely related to others, deposited in GenBank (Table 2 and Fig. 2), from India, Iran, France, Brazil, Singapore, Kenya, USA, and China. Similar to the P. aeruginosa gene, gene sequencing revealed a 99% similarity with other isolates deposited in GenBank.
Further, the E. coli blaVEB gene was closely related to those in isolates from France, USA, Norway, and Thailand and deposited in GenBank (Table 2 and Fig. 2). Again, gene sequencing revealed 99% compatibility with other Genbank-held genes. Furthermore, sequencing and BLAST analysis of the blaVEB genes isolated from the bacteria in this study showed the types of polymorphisms evident from the global standard genes (Table 3).
4. DISCUSSION
ESBLs are becoming significant causes of resistance to β-lactam antibiotics, especially in enteric bacteria such as K. pneumoniae, P. aeruginosa, and E. coli, resulting in severe consequences for the effective treatment of bacterial infections [13]. In the present study, the CLSI-recommended screening test detected 60 (92.3%) potential ESBL-producing isolates. However, the phenotypic method confirmed ESBL production in only 35 (58.3%) isolates. Ultimately, we were unable to confirm ESBL production in 25 isolates (41.6%). These results are likely due to the presence of other resistance mechanism(s), such as the presence of ambler Class C β-lactamases, as concluded in a previous study [14].
Multiple K. pneumoniae isolates were resistant to ceftriaxone (92.3%), ceftazidime (88.4%), aztreonam (80.7%), and cefepime but were generally more sensitive to the Carbapenems tested, similar to the results of Natoubi et al. [15]. Others have shown similar percentages of isolates being resistant to ceftazidime, ceftriaxone, and cefepime [16]. In the present study, K pneumoniae was the most frequent ESBL producing species, as also observed in another study [17]. However, these results contrast with those of some authors [18-20]who showed lower percentages of K. pneumoniae isolates being ESBL producers. These differences may be due to different degrees of exposure to β-lactam antibiotics.
Similar to the results obtained by Hakemi et al. [21], 34.2% of our P. aeruginosa isolates were ESBL-positive. Other authors [22, 23]reported lower rates of ESBL-positive P. aeruginosa isolates and one [24] reported a higher rate. These variations in ESBL-positive rates may be related to antibiotic use patterns in different geographic locations as well as differences in infection control procedures for hospital personnel in those regions. Moreover, the spread of ESBL-producing P. aeruginosa varies from country to country [22]. In the present study, Pseudomonas isolates showed resistance to ceftriaxone (72.4%), ceftazidime (65.5%), aztreonam (24.1%), and one was resistant to imipenem (3.4%), similar to the results by other investigators [22, 25].
Among the E. coli isolates included in the present study, high levels of resistance were observed against ceftriaxone (100%), ceftazidime (80%), aztreonam (100%), cefoxitin (90%), meropenem (20%), and imipenem (30%), similar to the rates reported by Hassan et al. [26]. In the present study, 14.4% of E. coli isolates were ESBL producers, similar to the reported rates of ESBL production in isolates in some studies,2 but much lower than in other studies [18, 19, 27].
Among the ESBL-producing isolates analyzed in the present study, 3 (8.5%) were carrying the blaVEB gene and 32 (91.4%) were not. This indicated that the majority of the isolates expressed ESBL activity encoded by a different gene, e.g., metallo-β-lactamase enzymes [28]. In this study, the prevalence of blaVEB genes was very low-one isolate in each of the three isolated Gram-negative species produced ESBLs. Among the ESBL-producing P. aeruginosa isolates, only 1 (8.5%) possessed the blaVEB gene. In contrast, two other investigations reported the prevalence of blaVEB genes to be 13.3% [29, 30]. In the current study, 1/18 (5.6%) ESBL-producing K. pneumoniae isolates possessed the blaVEB gene, whereas other studies have shown the prevalence of blaVEB genes among ESBL-producing K. pneumoniae to be 10.6% [31] to 12% [32]. In the present study, the blaVEB gene was present in 1/5 (20%) of ESBL-producing E. coli isolates. These results are much different from the results of other studies that showed the prevalence of the blaVEB gene in ESBL-producing E. coli to range from 0% [33-35] to 8% [36].
The phylogenetic trees of the blaVEB genes isolated in the present study were very similar to those isolated in other countries, with compatibilities of 99%, and showed high similarity with similar isolates from other countries.
CONCLUSION
Carbapenems (imipenem and meropenem) are the best choice for the treatment of ESBL producers of Gram-negative bacterial infections. The blaVEB gene plays an essential role in the resistance of ESBL-producing isolates to new β-lactam antibiotics. Further, the sequencing and phylogenetic analyses of the genes, from the P. aeruginosa, K. pneumonia, and E. coli isolates, revealed 99% similarity with the GenBank global standard genes.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
Not applicable.
HUMAN AND ANIMAL RIGHTS
No animals/humans were used for studies that are the basis of this research.
CONSENT FOR PUBLICATION
Not applicable.
AVAILABILITY OF DATA AND MATERIALS
The authors confirm that the data supporting the findings of this research are available within the article.
FUNDING
None.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
The authors acknowledge the English language editing assistance of Harkynn Consulting (www.harkynn.com).

