All published articles of this journal are available on ScienceDirect.

Prevalence of ESBL Genes Among Pseudomonas aeruginosa and Acinetobacter baumannii Isolated from Female UTI Patients in Dharwad, Karnataka, India
Authors Info & Affiliations
Abstract
Background
The emergence of ESBL-producing non-fermenters, such as Pseudomonas aeruginosa and Acinetobacter baumannii, is a growing concern, particularly in Urinary Tract Infections (UTIs). This study investigates the prevalence and distribution of ESBL genes among these pathogens isolated from female UTI patients in Dharwad, Karnataka, India.
Methods
A total of 713 urine samples were processed. Bacterial identification was conducted using culture and molecular methods. Phenotypic ESBL detection and quantitative PCR (qPCR) were employed to identify specific ESBL genes.
Results
Out of 713 samples, 665 were culture positive. Pseudomonas aeruginosa (56 isolates) and Acinetobacter baumannii (29 isolates) were identified. ESBL production was phenotypically confirmed in 69.6% and 48.3% of P. aeruginosa and A. baumannii, respectively. Genotypic screening revealed the high prevalence of blaSHV, blaTEM, and blaOXA-23 genes. blaKPC was only observed in P. aeruginosa.
Conclusion
The study demonstrates a high burden of ESBL-producing non-fermenters in UTI patients. Molecular surveillance is critical for effective antibiotic stewardship.
1. INTRODUCTION
Urinary Tract Infections (UTIs) remain one of the most common bacterial infections affecting individuals globally, with a significantly higher prevalence among women due to anatomical and physiological factors [1]. Although Escherichia coli is traditionally regarded as the predominant uropathogen, the role of non-fermenting Gram-negative bacilli (NFGNB), particularly Pseudomonas aeruginosa and Acinetobacter baumannii, in complicated and healthcare-associated UTIs is becoming increasingly significant [2, 3]. These opportunistic pathogens are associated with Multidrug Resistance (MDR), prolonged hospital stays, and increased morbidity and mortality. A major mechanism underlying antibiotic resistance in these pathogens is the production of Extended-Spectrum β-Lactamases (ESBLs), enzymes capable of hydrolyzing a wide range of β-lactam antibiotics, including penicillins, cephalosporins, and aztreonam [4]. While ESBL production is well documented in Enterobacteriaceae, there is growing evidence of ESBL gene dissemination among NFGNB, including P. aeruginosa and A. baumannii, largely mediated by mobile genetic elements such as integrons, plasmids, and transposons [5, 6].In the Indian context, the burden of ESBL-producing uropathogens is especially concerning due to high rates of antibiotic misuse, over-the-counter availability of antimicrobials, and suboptimal infection control practices [7]. Regional surveillance studies from various parts of India have reported alarming levels of MDR P. aeruginosa and A. baumannii isolates harboring ESBL genes such as blaTEM, blaSHV, and blaCTX-M [8, 9]. However, data on the prevalence of these resistance determinants among UTI isolates, particularly from female patients in Tier-II cities such as Dharwad, Karnataka, remain limited. Although Escherichia coli is considered the most prevalent uropathogen in community-acquired UTIs, Acinetobacter baumannii and Pseudomonas aeruginosa have emerged as clinically significant nosocomial pathogens, especially in complicated and catheter-associated UTIs. In immunocompromised and hospitalized patients, these organisms are frequently isolated [10]. They exhibit both intrinsic and acquired resistance mechanisms, particularly against β-lactams and carbapenems, which often results in ineffective treatment [11]. In addition, the World Health Organization (WHO) identifies P. aeruginosa and A. baumannii as critical-priority pathogens due to their ability to acquire and disseminate resistance genes via mobile genetic elements [12]. Given the critical public health implications of rising ESBL-mediated resistance, localized epidemiological studies are essential for guiding empirical therapy and implementing targeted antimicrobial stewardship strategies. This study aims to determine the prevalence of ESBL genes among P. aeruginosa and A. baumannii isolated from female UTI patients in Dharwad, Karnataka, India, and to assess their resistance patterns in the context of recent antimicrobial resistance trends.
2. MATERIALS AND METHODS
2.1. Study Design and Sample Collection
This prospective study was conducted between January and December 2023 at a tertiary care hospital in India. A total of 713 midstream urine samples were collected from female patients aged 16 to 70 years who were clinically suspected of having Urinary Tract Infections (UTIs). Sample size calculation was performed based on the prevalence of UTI in the Indian population. Samples were collected from both outpatients and hospitalized patients, as catheterization was considered one of the risk factors evaluated in the study. All samples were collected using sterile containers and processed within two hours of collection.
2.2. Isolation and Identification of Uropathogens
Urine samples were cultured on Blood agar, Luria-Bertani agar, Cysteine Lactose Electrolyte-Deficient (CLED) agar, and MacConkey agar using the calibrated loop streak method. Plates were incubated aerobically at 37°C for 24 hours. Significant bacteriuria was defined as growth of ≥10 5 CFU/mL. Isolates were identified using standard microbiological techniques and confirmed with the VITEK 2 Compact system (bioMérieux, France). Isolates were further subjected to molecular identification by 16S rRNA sequencing using the Sanger sequencing method.
2.3. Statistical Analysis
The association between various categorical risk factors and UTI positivity was analyzed using the Chi-square test of independence. P-values were calculated in GraphPad Prism version 10 (GraphPad Software, San Diego, CA, USA). A p-value <0.05 was considered statistically significant.
2.4. Antibiotic Susceptibility Testing
Antimicrobial susceptibility testing was performed by the Kirby-Bauer disk diffusion method on Mueller-Hinton agar, following Clinical and Laboratory Standards Institute (CLSI) guidelines [ 13 ]. The antibiotics tested included ampicillin (10µg), ciprofloxacin (30µg), imipenem (10µg), gentamicin (10µg), cefuroxime (30µg), ceftazidime (30µg), ceftriaxone (30µg), cefotaxime (30µg), cephalexin (30µg), cefepime (30µg), and nitrofurantoin (300µg). The isolates were distributed uniformly with a sterile, dry cotton swab on the Mueller-Hinton agar plate. Antibiotic discs were placed using sterile forceps. After incubating the plates at 37°C for 24 hrs, the results were interpreted according to Clinical Laboratory Standard Institute guidelines. For phenotypic detection of ESBL-producing isolates, isolates were selected based on resistance patterns to third-generation cephalosporins.
2.5. Screening of ESBL Producers
ESBL phenotypic detection was performed using the disc diffusion test. The Phenotypic Confirmatory Disc Diffusion Test (PCDDT) combines ceftazidime and clavulanic acid (30/10 µg), cefotaxime and clavulanic acid (30/10 µg), and ceftazidime (30 µg) and cefotaxime (30 µg) discs on Muller-Hinton agar with respective isolates. We kept the plates at 37 °C for 24 hours. An ESBL producer is defined as a zone diameter increase of greater than 5 mm for either ceftazidime/clavulanic acid disc or cefotaxime/clavulanic acid disc, compared to ceftazidime or cefotaxime alone.
2.6. Molecular Detection of Resistance Genes
Genomic DNA was extracted from isolates using a commercial DNA extraction kit. Quantitative PCR was used to detect ESBL and Carbapenemase genes, including blaTEM, blaSHV, blaGES, blaKPC, blaOXA-23, and blaOXA-48.The specific primer sets used for each gene are listed in Table 1. A loopful of bacterial isolates was suspended in 50 µL of ultrapure water.The suspension was centrifuged at 14000 rpm for 5 minutes, and the pellet was heated for 10 minutes at 95 °C to lyse the cells. The supernatant (30 µl) was collected and used as the DNA template for the reaction. qPCR was performed using the CFX96™ Real-Time PCR Detection System (Bio-Rad, USA). Each 20 µL reaction mixture contained 1 µL of DNA template, 0.8 µL each of forward and reverse primers (10 µM), 10 µL of 2× SYBR Green Master Mix (Bio-Rad), and nuclease-free water to make up the final volume. qPCR cycling conditions include Initial denaturation at 95 °C for 5 minutes, followed by 35 cycles of denaturation at 95 °C for 15 seconds and 60 °C for 30 seconds (combined annealing and extension), with fluorescence acquisition at the end of each cycle. A melt curve analysis was performed at the end of the run to confirm amplification specificity. qPCR was used to detect the presence or absence of the genes, and also ct values are qualitatively interpreted. All reactions were run in triplicate to ensure consistency and produce reliability of results. Klebsiella pneumoniae ATCC 700603 was used as a positive control for blaSHV, and for other genes, standard lab controls were used. Amplified products were visualized using gel electrophoresis, and melt curve analysis was performed to confirm gene specificity
3. RESULTS
3.1. Prevalence of Isolates and Patient Demographics
Out of 713 urine samples collected from female UTI patients, 665 (93.3%) yielded positive urine cultures. Among these, Pseudomonas aeruginosa was isolated in 56 cases (8.4%) and Acinetobacter baumannii in 29 cases (4.4%). The remaining isolates were predominantly Enterobacteriaceae (data not shown). The prevalence of P. aeruginosa and A. baumannii infections was highest among patients aged 41–50 years, followed by those aged 61–70 years, as illustrated in Fig. (1). The 16S rRNA gene sequences for representative P. aeruginosa (accession no. OQ363380) and A. baumannii (accession no. OQ363378) isolates were deposited in GenBank for reference. Among risk factors, only catheter use showed a statistically significant association (p < 0.05) with both species, suggesting it may be a significant risk factor for A. baumannii infections in our study (Table 2).
3.2. Antimicrobial Susceptibility Patterns
Both P. aeruginosa and A. baumannii isolates exhibited high levels of antimicrobial resistance, albeit with differing profiles. P. aeruginosa showed nearly universal resistance to certain first-line UTI drugs (notably 100% resistance to nitrofurantoin) and to third-generation cephalosporins (e.g., 80.3% of isolates were resistant to ceftazidime). However, this species retained relatively higher susceptibility to a few antibiotics: only 16.1% of P. aeruginosa isolates were resistant to ciprofloxacin, followed by low resistance rates to gentamicin (21.4%) and imipenem (23.2%). Resistance to cefotaxime was moderate (observed in 57.2% of isolates). In contrast, A. baumannii demonstrated extreme drug resistance to many tested agents. Notably, 75% of A. baumannii isolates were resistant to ampicillin, and similarly high resistance frequencies were observed for the oral cephalosporin cefalexin and the parenteral cephalosporins ceftazidime and ceftriaxone (each with over half of the isolates being non-susceptible). All A. baumannii isolates were also resistant to nitrofurantoin. These antimicrobial resistance patterns for both organisms are summarized in Table 3.
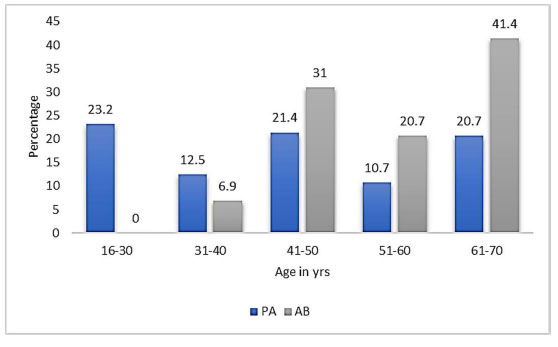
Prevalence of uropathogens in female patients with UTI.
| Genes | Primer Sequence (5’ - 3’) FW: Forward RV: Reverse |
Product Size (bp) | Annealing Temp (°C) |
|---|---|---|---|
| blaTEM | FW: GTATCCGCTCATGAGACAATAACC RV: CCAATGCTTAATCAGTGAGGCACC |
918 | 42.5 |
| blaSHV | FW: CGCCTGTGTATTATCTCCCTG RV: TTGCCAGTGCTCGATCAGCG |
842 | 44.5 |
| blaGES | FW: CTATTACTGGCAGGGATCG RV: CCTCTCAATGGTGTGGGT |
594 | 51 |
| blaKPC | FW: CATTCAAGGGCTTTCTTGCTGC RV: ACGACGGCATAGTCATTTGC |
538 | 51 |
| blaOXA-23 | FW: GTGGTTGCTTCTCTTTTTCT RV: ATTTCTGACCGCATTTCCAT |
736 | 50.3 |
| blaOXA-448 | FW: AACGGGCGAACCAAGCATTTT RV: TGAGCACTTCTTTTGTGATGGCT |
585 | 50.3 |
| Risk factors | Frequency (%) | |
|---|---|---|
| P.aeruginosa | A. baumanii | |
| Cathetar use | 8 (16.1) | 14 (48.3) |
| Menopause | 13 (23.2) | 11 (37.9) |
| History of UTI | 12 (21.4) | 5 (17.2) |
| Diabetes | 9 (16.1) | 8 (27.6) |
| Sexual activity | 8 (16.1) | 7 (24.1) |
3.3. Phenotypic Detection of ESBL Production
Phenotypic screening tests identified a subset of the isolates as Extended-Spectrum Β-Lactamase (ESBL) producers. Among the 56 P. aeruginosa isolates, 39 (69.6%) were confirmed to be ESBL producers, and among the 29 A. baumannii isolates, 14 (48.3%) were ESBL-positive. Supplementary Fig. (1) shows a representative phenotypic confirmatory test for ESBL production in these organisms. All ESBL-producing P. aeruginosa and A. baumannii isolates were selected for further molecular characterization.
| Genotype | P. aeruginosa (n=39) | A. baumanii (n=14) |
|---|---|---|
| blaTEM | 27 (69.2) | 9 (64.3) |
| blaSHV | 33 (84.6) | 8 (57.1) |
| blaGES | 11(28.2) | 6 (42.8) |
| blaKPC | 19 (48.7) | - |
| blaOXA-23 | 32 (82) | 11 (78.6) |
| blaOXA-48 | 28 (71.8) | 5 (35.7) |
| blaTEM + SHV | 13 (33.3) | 7 (50) |
| blaOXA-23 + OXA-48 | 9(23.1) | - |
3.4. Genotypic Characterization of ESBL Genes by qPCR
All phenotypic ESBL-positive isolates were subjected to quantitative PCR analysis targeting ESBLs and carbapenemase genes (blaTEM, blaSHV, blaGES, blaKPC, blaOXA-23, and blaOXA-48). Genotypic results revealed that in P. aeruginosa (n = 39), the most prevalent ESBL gene was blaSHV, detected in 84.6% of isolates, followed by blaTEM (69.2%) and blaGES (28.2%). The blaKPC gene was also detected in P. aeruginosa (in nearly half of the ESBL-positive P. aeruginosa isolates), whereas blaKPC was not found in any A. baumannii isolate. In A. baumannii (n = 14), the carbapenemase gene blaOXA-23 was the most common, present in 78.6% of isolates, and blaSHV was identified in 57.1%. The blaOXA-48 gene appeared at a lower frequency in A. baumannii (detected in roughly one-third of isolates) compared to P. aeruginosa. Notably, blaTEM was also frequently detected in A. baumannii, often co-occurring with blaSHV in the same strains. In our collection, neither species showed a high prevalence of the blaGES gene in A. baumannii (this gene was either absent or present at very low frequency in the A. baumannii isolates). Multiple ESBL genes were common among the tested strains. One-third (33.3%) of the ESBL-producing P. aeruginosa carried more than two of the targeted resistance genes concurrently, and fully half (50%) of the ESBL-producing A. baumannii harbored at least two or more ESBL genes (often including both blaTEM and blaSHV). The detailed prevalence of each bla gene and its co-occurrence in these isolates is presented in Table 3. Cycle threshold (Ct) values for the amplification of each gene target (indicative of gene abundance) are provided in Supplementary Table 1. Finally, the qPCR amplification and melt curves confirming the specific gene amplifications are shown in Fig. (2) demonstrating successful amplification of the target ESBL genes in the representative P. aeruginosa and A. baumannii samples
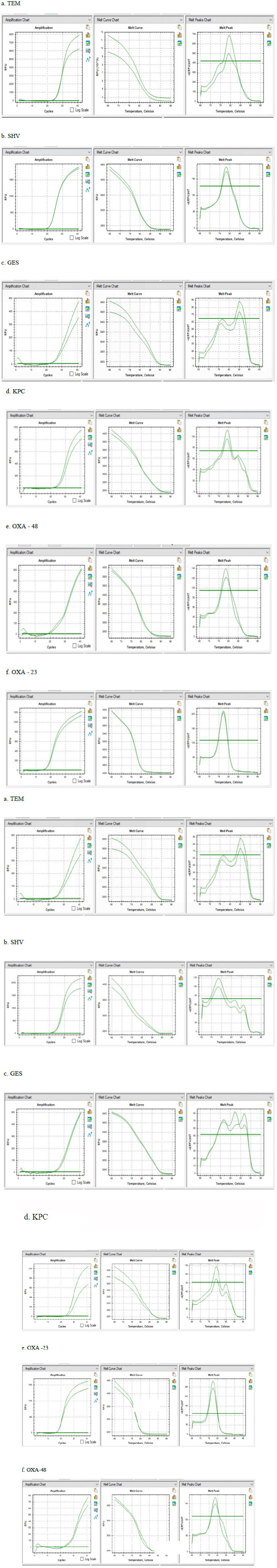
PCR amplification and melt peak curves of genes in respective uropathogens.
4. DISCUSSION
The rise of Multidrug-Resistant (MDR) uropathogens, particularly among non-fermenting Gram-negative bacteria such as P. aeruginosa and A. baumannii, represents a significant clinical challenge both globally and within hospital settings in India. This study reveals a substantial prevalence of ESBL-producing P. aeruginosa and A. baumannii among female UTI patients in Dharwad, Karnataka, highlighting both organisms as critical nosocomial threats. Notably, ESBL production was phenotypically confirmed in 69.6% of P. aeruginosa and 48.3% of A. baumannii isolates, a pattern consistent with previous reports suggesting increasing resistance among non-fermenters [5, 14].
The high proportion of isolates harboring ESBL genes, particularly blaSHV(84.6%) and blaTEM (69.2%) in P. aeruginosa, underscores the genetic adaptability and resistance potential of this pathogen. Our detection rates exceed those reported in similar studies from the Middle East and South Asia, which found blaSHV and blaTEM in fewer than 50% of clinical Pseudomonas isolates [15, 16]. In A. baumannii, the predominance of blaOXA-23 (78.6%) aligns with findings from Algeria and Iran, where this gene is frequently associated with carbapenem resistance [17, 18]. The detection of blaOXA-48 in 71.8% of P. aeruginosa and 35.7% of A. baumannii is alarming and represents a potential reservoir for pan-drug resistance, as OXA-48-like enzymes contribute to carbapenem hydrolysis while remaining undetectable by many phenotypic tests [19]. Of concern, nearly one-third of P. aeruginosa isolates and half of A. baumannii isolates harbored two or more ESBL genes, suggesting a strong potential for horizontal gene transfer and co-selection of resistance traits. Such gene combinations may facilitate survival under a broad spectrum of antibiotic classes, especially in hospital environments where antimicrobial pressure is high [20].
Furthermore, greater resistance to nitrofurantoin and high resistance to β-lactams and cephalosporins, particularly to ceftazidime and cefotaxime, further reinforce the growing clinical irrelevance of these agents for empirical UTI therapy. Compared to recent data from Nepal and Poland, where ceftazidime resistance in P. aeruginosa ranged between 40–60%, our observed resistance rate of 80.3% suggests region-specific antimicrobial misuse or selective pressure [21, 22]. In contrast, relatively preserved susceptibility to imipenem, ciprofloxacin, and gentamicin (resistance <25%) indicates their potential role as second-line therapies, although this should be approached cautiously due to the risk of emerging carbapenem resistance [23].
Hormonal factors, such as menopause, were associated with higher rates of isolation of P. aeruginosa and A. baumannii, echoing the existing literature linking estrogen deficiency to changes in the genitourinary flora and increased infection risk [24]. The role of urinary catheterization as a prominent risk factor for A. baumannii infections (48.3%) further underscores the organism’s opportunistic nature and its propensity to form biofilms on indwelling devices [25].
Our study demonstrates the utility of qPCR as a rapid and sensitive method for detecting ESBL genes, with Ct values indicating robust amplification efficiency across most targets. The use of melt curve analysis provided additional specificity in distinguishing amplicon identities. Taken together, these findings reinforce global and regional alerts on AMR, as outlined in the WHO priority pathogen list [26]. The presence of high-risk clones carrying multiple β-lactamase genes necessitates ongoing molecular surveillance and stringent infection control practices. Moreover, our results suggest the urgent need to revise empirical UTI treatment protocols in this region, incorporating local resistance data to avoid therapeutic failures and reduce selective pressure.
CONCLUSION
This study indicates that Pseudomonas aeruginosa and Acinetobacter baumannii showed high incidence of resistance genes bla TEM , bla SHV , and bla OXA-23. These uropathogens represent a wide range of clinical complications due to their high rates of multidrug resistance, co-occurrence of other resistance genes, and significant association with risk factors like catheterization. Although antibiotics such as nitrofurantoin, gentamicin, ampicillin, and cefotaxime are not recommended against these uropathogens, we evaluated them in our study due to their empirical use by clinicians in our study setting, which further highlights the implications of local prescribing patterns. The use of qPCR has enhanced its importance in monitoring antibiotic resistance by enabling quick, precise identification of ESBL and carbapenemase genes. These findings emphasize the critical need for antimicrobial stewardship, routine molecular diagnostics, and infection control practices. Continuous surveillance and targeted antimicrobial therapy to manage and control the spread of resistant strains are necessary.
LIMITATION
This study has some limitations. The limited number of isolates of P. aeruginosa and A. baumannii may affect generalizability. Resource constraints hindered the incorporation of the bla CTX-M gene, even though they have worldwide significance. Resistance mechanisms such as efflux pumps and porin loss were not investigated. Molecular typing methods are important to study clone-relatedness and transmission of resistant strains, this will be considered in future studies. In addition, the study is limited by its single-center design and sequencing to determine the genetic context of the ESBL genes.
AUTHORS' CONTRIBUTIONS
The authors confirm their contributions to this paper as follows. CU was responsible for the study conception and design. MH and RG contributed to conceptualization. RN and RM were responsible for data curation, and NB conducted the investigation. All authors reviewed the results and approved the final version of the manuscript.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
The study was approved by the KLES Kidney Foundation Institutional Ethics Committee, Belagavi, with approval Number KLESKF/ IEC/23/017.
HUMAN AND ANIMAL RIGHTS
No animals were used in this research. All procedures performed in studies involving human participants were in accordance with the ethical standards of institutional and/or research committee and with the 1975 Declaration of Helsinki, as revised in 2013.
AVAILABILITY OF DATA AND MATERIALS
The data and supportive information are available within the article.
ACKNOWLEDGEMENTS
We express our gratitude to CSIR NET UGC for providing financial support in the form of a JRF, under the Roll number KK10000775, and to other laboratories for providing clinical samples. The study received no funding from any agency.

