All published articles of this journal are available on ScienceDirect.

Unravelling the Probiotic and Safety Profile of Lactiplantibacillus plantarum 022AE: A Multi-Omics Approach Integrating Genomics and Phenotypic Data
Authors Info & Affiliations
Abstract
Introduction
Lactiplantibacillus plantarum is one of the most varied species of lactic acid bacteria in various environments. Probiotics are beneficial organisms that help balance the gut microbiome and promote general health. The purpose of this study was to perform preliminary phenotypic and genetic characterization of the probiotic strain L. plantarum 022AE.
Method
Using hybrid assembly, L. plantarum 022AE was sequenced, producing a 3.23 Mb scaffold. Using NCBI-BLASTN, the strain's identification was verified. Genome annotation was used to evaluate safety characteristics. In vitro tests were employed to assess cytotoxicity, antibacterial activity, bile acid tolerance, epithelial adhesion, surface characteristics, cell lining to verify in vivo adhesion, and antibiotic susceptibility (CLSI). Genes associated with virulence, adhesion, and stress survival were discovered by comparative genomics. The safety and probiotic qualities of L. plantarum 022AE were shown to be favourable by full-genome analysis and phenotypic assessment.
Results
For complete genome sequencing of L. plantarum 022AE, a single scaffold of 3,234,271 bp was obtained via hybrid assembly. NCBI-BLASTN tools analysis showed L. plantarum 022AE to be 100% identical to the reference strain HAC01. Gene annotation and downstream analysis revealed safety attributes, like absence of transferable antibiotic resistance genes, virulence factor genes, active biogenic amine-producing genes, enterotoxin genes, emetic toxin genes, and prophage sequences. In vitro phenotypic characterization showed that the strain was bile- and acid-tolerant, attached to intestinal epithelial cells, and exhibited favorable cell surface properties and antimicrobial activity against key pathogens. It was susceptible to CLSI-recommended antibiotics and produced no cytotoxicity or enterotoxicity. Comparative genome analysis of L. plantarum 022AE revealed genetic determinants for survival in stress environments, cell adhesion, and virulence factor genes.
Conclusion
The entire genome study and phenotypic evaluation exhibited a positive profile in terms of safety and probiotic attributes of L. plantarum 022AE, in alignment with current regulatory standards, suggesting its potential for use in applications requiring safe microbial strains, particularly in the food and pharmaceutical industries.
1. INTRODUCTION
Lactiplantibacillus plantarum is a gram-positive, non-motile lactic acid bacterium (LAB) that lives in mesophilic and microaerophilic environments and produces no spores. It is one of the most adaptable LAB species, as evidenced by its capacity to colonize a diverse range of habitats, such as the gastrointestinal and vaginal tracts, vegetables, dairy products, and fermented foods. It is extensively utilized in industrial fermentation due to its “generally recognized as safe” (GRAS) and qualified presumption of safety (QPS) status [1]. The proven functional and health-promoting qualities of L. plantarum strains have bolstered interest in their uses during the past century [2]. L. plantarum has a wide range of beneficial qualities, making it the most popular probiotic. These include its ability to decrease cholesterol levels, alleviate lactose intolerance, improve the intestinal barrier, and alter the commensal microbiota [3]. Specifically, the 022AE strain examined in this paper has demonstrated potential advantages in multiple investigations, including the ability to survive passage through the human gastrointestinal tract, suitable aggregation properties, production of antioxidants, GI tract adherence potential, and anti-microbial compound producing capabilities, as assessed in in vitro assays [3].
All microorganisms purposefully utilized in the food chain must undergo an unambiguous taxonomic identification at the strain level, according to the EFSA's statement and the FEEDAP guidance document [2, 4]. Strains can differ significantly from one another, even within the same species, and the traits attributed to one strain may not always be applicable to another [2].
The characterization of each probiotic strain thus requires the use of whole-genome sequencing (WGS) data and WGS-based data processing. In addition to information regarding virulence factors, antibiotic resistance, and the generation of hazardous compounds, WGS can offer important insights into the characterization of these strains' putative functional characteristics. In the past, many potential probiotics strains have been identified and characterized by the WGS [5]. Genomic analysis of L. plantarum 022AE revealed that it harbors many genes, which contribute to its probiotic properties, as well as lacks genes, which might pose safety concerns.
This comprehensive study, using a dual approach of WGS-based genomic properties and phenotypic assays of L. plantarum 022AE, has provided extensive data to explore its probiotic potential.
2. MATERIALS AND METHODS
2.1. Bacterial Strains, Media, Chemicals, and Genomic DNA Extraction and Purification
The L. plantarum 022AE strain used in this experiment was obtained from an in-house proprietary technique at Advanced Enzymes Technology Ltd. Table 1 lists pathogenic bacterial and yeast strains, as well as the conditions under which they flourish. All reagents and chemicals were obtained from Sigma Aldrich, India, and microbiological media were obtained from Hi Media Labs Pvt. Ltd., India. Nanodrop-2000, Qubit, and agarose gel electrophoresis were used to determine the quantity and quality of genomic DNA from the L. plantarum strain 022AE [3, 6]. The DNA yield and purity (260/230 and 260/280 ratios) were enough for creating an NGS library [7].
| Sr. No. | Pathogenic Bacteria | Growth Medium Used | Assay Medium Used |
|---|---|---|---|
| 1. | Bacillus cereus ATCC 33019 | Nutrient broth | Mueller-Hinton agar |
| 2. | Bacillus circulans ATCC 4516 | Nutrient broth | Mueller-Hinton agar |
| 3. | Bacillus subtilis subsp. spizizenii ATCC 6633 | Brain heart infusion broth | Mueller-Hinton agar |
| 4. | Candida albicans ATCC 90028 | Potato dextrose broth | Mueller-Hinton agar |
| 5. | Clostridium difficile ATCC 9689 | Reinforced Clostridial medium broth | Reinforced Clostridial medium agar |
| 6. | Clostridium perfringens ATCC 13124 | Reinforced Clostridial medium broth | Reinforced Clostridial medium agar |
| 7. | Clostridium sporogenes NCIM-5125 (equivalent to ATCC 19404) | Reinforced Clostridial medium broth | Mueller-Hinton agar |
| 8. | Enterobacter cloacae ATCC 13047 | Nutrient broth | Mueller-Hinton agar |
| 9. | Escherichia coli ATCC 700728 | Nutrient broth | Mueller-Hinton agar |
| 10. | Escherichia coli ATCC 9002 NCTC | Nutrient broth | Mueller-Hinton agar |
| 11. | Klebsiella pneumoniae ATCC BAA-1144 | Soybean casein digest broth | Mueller-Hinton agar |
| 12. | Listeria monocytogenes ATCC 19115 | Brain heart infusion broth | Brain heart infusion agar |
| 13. | Micrococcus luteus MTCC 106T | Brain heart infusion broth | Mueller-Hinton agar |
| 14. | Pasteurella multocida ATCC 12945 | Brain heart infusion broth | Tryptone soy agar |
| 15. | Pseudomonas aeruginosa ATCC 9027 | Nutrient broth | Mueller-Hinton agar |
| 16. | Salmonella abony NCIM-2257(Equivalent to ATCC 6017 NCTC) | Nutrient broth | Mueller-Hinton agar |
| 17. | Salmonella enterica ATCC 14028 | Nutrient broth | Mueller-Hinton agar |
| 18. | Staphylococcus aureus ATCC 6538P | Nutrient broth | Mueller-Hinton agar |
2.2. Whole Genome Sequencing
Genomic DNA from L. plantarum strain 022AE was used for Illumina and Nanopore whole-genome sequencing (WGS). The Sure Select QXT kit was used to fragment DNA, tag it with an adaptor, and purify it for Illumina sequencing. Following 6-cycle PCR amplification, the library was purified, quality-checked, and assessed for fragment size (200-700 bp). The Illumina MiSeq platform generated 2,941,678 reads. To carry out Nanopore sequencing, one µg of DNA was end-repaired, barcoded, and pooled at equivalent amounts before adapter ligation. The Grid ION X5 sequenced the final library using a 48-hour procedure. The raw readings were base-called and processed with MaSuRCA for hybrid genome assembly. The genome was constructed and annotated utilizing the NCBI Prokaryotic Genome Annotation Pipeline (PGAP) [8-10].
2.3. Phenotypic Characterization
Phenotypic characteristics of Lactobacillus plantarum 022AE have been studied and reported by Namrata Bhingardeve et al. The strain was tested for stability at various pH levels (1.5–7.0) and bile concentrations (0.01–1.0%) at 37°C. The pour plate method was used to determine viability every hour for 5 hours. To investigate the stability of L. plantarum 022AE in a simulated gut model, L. plantarum 022AE was introduced to several food matrices and examined under simulated digestion circumstances (salivary, gastric, and intestinal fluids) using the COST INFOGEST protocols [3]. Viability was assessed after each stage. An extensive study was conducted using fasting and fed conditions. Hydrophobicity was measured using the bacterial adhesion to hydrocarbons (BATH) method, and auto-aggregation was evaluated by detecting cell clumping over 6 hours. L. plantarum 022AE was mixed with pathogens, and their co-aggregation ability was determined by measuring OD600 at 0 and 6 hours [3]. The adherence of bacteria to mucin was investigated using a microplate assay. After incubation, non-adherent cells were washed away, and live bacteria were counted on MRS agar. ONPG broth's color shift confirmed β-galactosidase activity. Precipitation around colonies cultured on bile salt agar revealed bile salt hydrolase (BSH) activity [3]. The DPPH radical scavenging activity was assessed in a microplate experiment, and the antioxidant potential was calculated using absorbance data. Antimicrobial compounds (AMCs) were isolated from L. plantarum 022AE with XAD16N beads and evaluated against 18 diseases using the spot-on-the-lawn method [3]. Zones of inhibition were recorded. L. plantarum 022AE stability was examined in a variety of liquid matrices (water, buffer, oil, and emulsions) at both real-time (5°C) and accelerated conditions (25°C). Thermal stability was tested for 6 hours at temperatures ranging from 4 to 50 degrees Celsius. All experiments were carried out in three replicates. The data were analyzed using GraphPad Prism with ANOVA and multiple comparison tests (Tukey's HSD or Dunnett's). A two-tailed Student's t-test was employed to evaluate mucin adhesion [3, 11].
2.4. Technological Characterization
Human colon adenocarcinoma cell line (Caco-2) cells were cultured in minimum essential media (MEM) with 20% fetal bovine serum at 37ºC and 5% CO2. Media was refreshed every 2-3 days. Caco-2 cells (1×105 cells/ml) were seeded in six-well plates and incubated at 37°C and 5% CO2 [12]. Until the cells achieved 80% confluency, the media was changed every 48 hours. MEM was used to replace spent medium (antibiotic-free), which was incubated at 37°C for 30 minutes [6]. The cells were then washed twice with PBS (pH 7.4). One mL of serum- and antibiotic-free MEM was added, and the mixture was incubated at 37 °C for 30 minutes [6]. Bacterial isolates (1×109 CFU in 1 ml MEM) were then added to wells. Plates were incubated for two hours at 37°C with 5% CO2. The monolayer was washed five times with PBS to remove non-adherent bacteria. The cells were then fixed with 2 ml methanol for 10 minutes and stained with 3 ml Giemsa (1:20 in PBS) for 20 minutes. They were afterward rinsed with distilled water, air-dried, and examined under a 40X microscope. The bacteria were counted in 20 random fields, and adhesion was classified as non-adhesive (≤40), adhesive (41–100), or strongly adhesive (>100) [13, 14]. To measure the percent adhesion, the monolayer was washed five times with PBS to remove non-adherent bacteria. Cells were then detached by incubating with 1 mL of 0.25% trypsin-EDTA for 15 minutes [14]. Cell bacteria were then diluted in saline suspension and plated on MRS agar. Viable bacteria were then enumerated after incubation. The adhesion was calculated using formula (1) as follows:
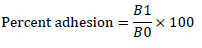 |
(1) |
Where, B0 and B1 are initial and final CFU counts.
2.5. Safety Assessment
2.5.1. Cytotoxic Activity Of Bacterial Culture Supernatants Using Vero Cells
A cytotoxicity assay using Vero cells was performed to demonstrate if L. plantarum 022AE is free of toxigenic potential [15]. The test is based on the principle that the DNA intercalating agent propidium iodide will stain DNA in cells with leaky membranes, thereby enhancing the resulting intracellular fluorescent signal. The DNA of intact cells would not show any uptake of propidium iodide, resulting in a basal level of negligible fluorescence. The positive control Triton X-100-treated cells with leaky cell membranes show 100% fluorescence. A compound is considered active if the fluorescence unit (FU) values of the test sample are 20% or above compared to the values obtained with the positive controls (100%) [16]. The cytotoxicity of L. plantarum 022AE was measured in triplicate at three different concentrations [17].
2.5.2. Detection Of Enterotoxins in Lactobacillus plantarum 022AE
The presence of enterotoxins in L. plantarum 022AE was tested using the Duopath® enterotoxin test kit. Positive controls were Bacillus cereus ATCC 14579 and Bacillus cereus ATCC 11778. It was observed that L. plantarum 022AE did not produce enterotoxins, while the toxin was detected in positive control strains [18, 19]. L. plantarum 022AE was inoculated into Lactobacillus MRS broth (loopful culture). To prepare the sample, 10 mL of inoculum was combined with 90 ml of CGY broth (1% glucose) and homogenized with a stomacher. The mixture was incubated at 37 °C for 18-24 hours. After enrichment, 200 µL of the sample was placed into 20 ml of new CGY broth (with 1% glucose) in a 200 mL flask and incubated for 6 hours at 37°C with shaking (120 rpm) [20]. Prior to testing, the enriched sample and test device were brought to room temperature (15-25°C). The test device was put on a flat table and correctly tagged. Testing was carried out within two hours of the device being opened. A 150 µL-processed sample was pipetted into the test device's circular sample port and incubated at room temperature [21]. The test result was obtained 30 minutes after the sample was administered. A red line on ‘C’ confirmed the device to be functioning properly. If a red line appeared only at zone ‘C’, the test was negative. The test was positive if a red line appeared in the ‘NHE’ and/or ‘HBL’ zone along with zone ‘C’ [18, 19].
2.5.3. Antibiotic Susceptibility Testing
As advised by EFSA (2012), the antibiotic susceptibility of L. plantarum strain 022AE was evaluated in accordance with CLSI (2012, 2016) recommendations [22-24]. The sensitivity of L. plantarum 022AE to nine antibiotics, recommended by CLSI, was assessed. Streptococcus pneumoniae ATCC 49619 was employed as the control strain for the microbiological tests. The tested antibiotic's minimum inhibitory concentration (g/ml) against L. plantarum strain 022AE fell within the EFSA (2012) recommended breakpoints [22]]. The minimum inhibitory concentrations (MICs) of antibiotics that inhibited the growth were identified. EFSA (2012) and CLSI (2015) breakpoints were used to classify the results as either “S” (susceptible) or “R” (resistant). The results obtained were the average of three replicates, as the assay was run in triplicate [23].
2.5.4. Genomic Characterization
Following the method provided by Salvetti et al.. (2016), the assembled genome's stability was evaluated. Investigations were conducted on CRISPR, prophage, and insertion sequences (IS). PHASTER was used to identify prophage sequences, while ISfinder and the ACLAME database (version 0.4) were employed to detect mobile genetic elements. CRISPRCasFinder was used to screen for CRISPR sequences [25]. The assembled genome of L. plantarum strain 022AE was assessed for genomic features related to probiotic properties, including adhesion to gut mucosa, acid tolerance, bile salt tolerance, and environmental stress resistance [25]. Predicted protein sequences were annotated using the Batch-CD Search web service in the Conserved Domain Database, with the Pfam database (containing 19,178 position-specific scoring matrices) selected for functional annotation. A search for similarities between the assembled genome of L. plantarum strain 022AE and the Comprehensive Antibiotic Resistance Database (CARD) was performed using BLASTX with the following criteria: similarity >30%, coverage > 70%, and e-value < 1e-02, to identify significant hits [16, 26]. Virulence factor genes/proteins were downloaded from the VFDB (Virulence Factor Database), containing 3072 sequences [27]. A search for similarities between the assembled genome of L. plantarum strain 022AE and virulence factor proteins was performed using BLASTX with the following criteria: similarity >30%, coverage > 70%, and e-value < 1e-02, to identify significant hits. In accordance with Andrea and Salvetti, BLASTX was used to explore the genome of L. plantarum strain 022AE for genes linked to biogenic amine synthesis, primarily amino acid decarboxylases. BLASTX was used to compare the assembled genome with the protein sequences of biogenic amine-producing genes (amino acid decarboxylases) that were obtained from the UniProt database [16, 25].
2.5.5. Comparative Genomics
The assembled genome of L. plantarum strain 022AE was compared with other bacterial genomes present in the RefSeq genome database using NCBI-BLASTN [16]. High-quality processed reads were mapped against the closest reference genome (L. plantarum strain HAC01) using BWA -v0.7.17 to find out the mapping statistics [28]. Using the online ANI calculator tool, average nucleotide identity (ANI) analysis was carried out between the assembled genome of L. plantarum strain 022AE and the reference genome L. plantarum strain HAC01. A synteny plot between the assembled genome of L. plantarum strain 022AE and reference (L. plantarum strain HAC01) was generated using Mauve version 2.4.0 based on default parameters. Mauve performs genome alignment to identify evolutionary changes in the DNA by aligning homologous regions of sequence. Circular genome comparison between the two genomes was carried out using BRIG-V0.95 based on default parameters [29, 30].
3. RESULTS
3.1. Genome Assembly of L. plantarum 022AE
A total of 272,704 reads with an average length of 3,651 bp were generated using a nanopore sequencer, yielding 995 Mb bases with >300× genome coverage. The final genome assembly was 3,234,271 bp with a GC content of 44.55% and was deposited in the NCBI/GenBank database (accession: CP031127). Gene annotation was performed using the UniProt database (52,531 proteins for Lactobacillus plantarum). Of the 3,047 coding sequences (CDS) identified, 3,035 showed significant matches (>30% identity, e-value ≤1e-05) with UniProt Lactobacillus proteins. Gene ontology analysis classified these proteins into molecular function (40.11%), cellular component (44.12%), and biological process (15.31%), as shown in Figs. (1-3).
3.2. Phenotypic Characterization
Namrata Bhingardeve et al. [3] studied the phenotypic characteristics of L. plantarum 022AE, and each experiment was carried out in triplicate; the results were expressed as mean ± standard deviation (SD) in Log10 CFU/g or mL. Statistical tests and graph preparation were performed using GraphPad Prism (v8.0.2; GraphPad Software Inc., USA; https://www.graphpad.com/scientifc- sofware/prism/). Differences between groups were assessed using two-way ANOVA, followed by either Tukey’s HSD or Dunnett’s multiple comparison test, with significance considered at p < 0.05. The strain survived in an acidic environment for up to 5 hours at pH 3.5–7.0 without any noticeable loss. The strain was able to withstand bile concentrations of up to 1% for 3 hours; however, after 4 hours, it began to slightly decline. In a simulated gut model, free cells survived all digestion phases, and there was no noticeable change in gastric or intestinal conditions. Viability was either maintained or improved in the presence of food matrices (milk, baby food, SAD, and SED), especially in the intestinal phase. The highest adherence to ethyl acetate (25.2%) and a moderate amount of autoaggregation (16.13%) were shown by L. plantarum 022AE. L. plantarum 022AE co-aggregated with S. aureus (14.75%) and Candida albicans (20.45%). Bile salt hydrolase activity and β-galactosidase production were examples of functional characteristics. Moderate antioxidant activity of cell-free extracts was demonstrated by 24.61% DPPH scavenging activity. In in vitro tests, antimicrobial activity was observed against 15 out of 18 pathogenic bacteria. The stability of L. plantarum 022AE cells was maintained at temperatures of 4 to 50°C. The strain was viable for 6 hours at 4 to 25°C, for 5 hours at 40°C, and for 2 hours at 50°C. L. plantarum 022AE maintained viability in McIlvaine buffer (97.8%) for 6 months. Statistical analysis is presented in the supplementary data sheet.
3.3. Technological Characterization
Cell adhesion potential of L. plantarum 022AE was evaluated on human epithelial cells (Caco-2 cells). Cell adhesion potential was evaluated using two methods, i.e., observation of bacterial adhesion under microscope (adhesion score) and counting the adhered cell colonies after trypsinization (percent adhesion calculation) (Fig. 4). Based on observations of direct adhesion of bacteria to Caco-2 cells, adhesion score of L. plantarum 022AE was 160.55±16.81, indicating strong adherence to human epithelia cells (Caco-2 cells). Table 2 presents the percent adhesion data.
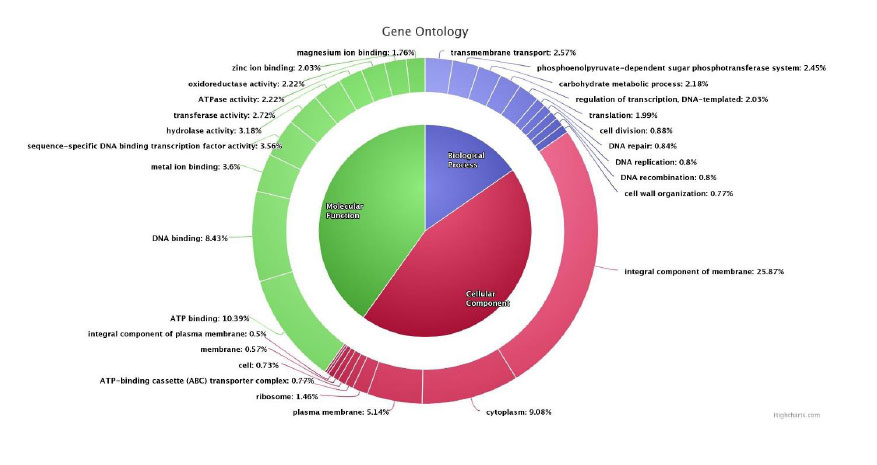
Gene ontology (GO) association of predicted protein coding genes (molecular function – 40.11%, cellular components – 44.12%, and biological process – 15.31%).
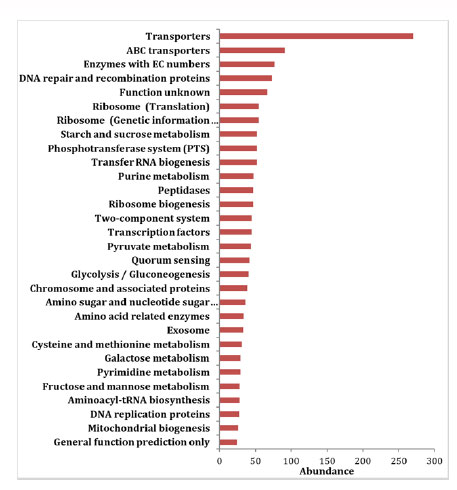
Pathway abundance of predicted proteins.
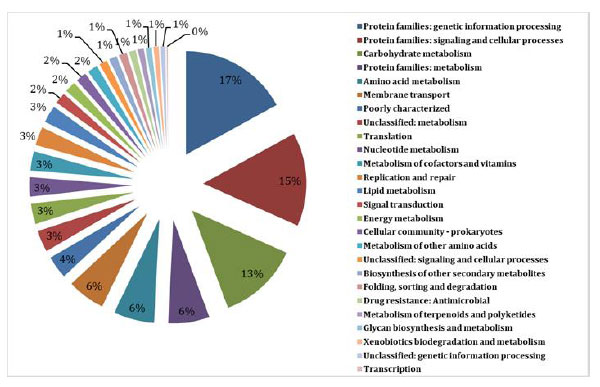
Pathway function associated with predicted proteins.
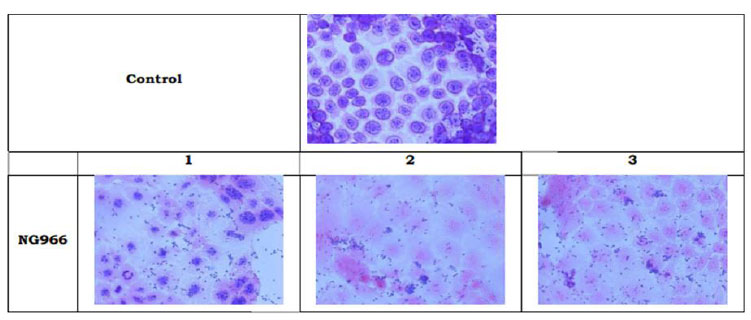
Adhesion of bacterial cultures on Caco-2 cell culture.
| 022AE | Adhesion Score | Adhesion Percentage | Adhesion Category |
|---|---|---|---|
| 160.55±16.81 | 7.97±0.54 | Adhesive |
3.4. Safety Assessment
3.4.1. Cytotoxic Activity of Bacterial Culture Supernatants using Vero Cells
The fluorescence values observed for the Lactobacillus plantarum 022AE sample were less than 20% of the positive control, indicating that the sample did not exert any cytotoxic effects in vitro. Vero cells were incubated with 10-100 µL of the sample for 2 hours (Table 3).
3.4.2. Detection Of Enterotoxins in Lactobacillus plantarum 022AE
All experimental devices showed a red line zone ‘C’, which indicates the functioning of the test device. The test microbe L. plantarum 022AE showed no red line in zones NHE or HBL, which indicated the negative test result, i.e., absence of both NHE and HBL enterotoxins. Control samples B. cereus ATCC 14579 and B. cereus ATCC 11778 showed a red line in NHE along with a red line in zone ‘C’, which indicated the positive test result, i.e., the presence of NHE enterotoxins (Fig. 5A-C).
3.5. Antibiotic Susceptibility Testing
L. plantarum strain 022AE was tested as per CLSI guidelines for its susceptibility/resistance against antibiotics, viz., clindamycin, chloramphenicol, ampicillin, gentamicin, tetracycline, kanamycin, vancomycin, and erythromycin. L. plantarum strain 022AE was sensitive to all tested antibiotics (Table 4). The MIC breakpoint values observed for L. plantarum 022AE were well below or equal to the breakpoint values described by EFSA (2012).
3.6. Genomic Characterization
3.6.1. Antibiotic Resistance Genes And Safety Analysis
The genome of L. plantarum strain 022AE showed 347 possible antibiotic resistance genes related to several functional categories, including signal transduction, transcription, defence mechanisms, and metabolism. Important functional areas included transcription (130 genes), defence mechanisms (72 genes), and cell wall/membrane formation (21 genes). Further screening against critically essential antimicrobials, as defined by WHO (2016) and EFSA (2012), provided just three full-length coding genes (Table 5). These genes have been found to be essential to the species and not associated with resistance acquisition.
These resistant genes’ surrounding areas were analyzed using the ISfinder and ACLAME databases, which verified that there was no possibility of horizontal gene transfer due to the lack of mobile genetic elements. Following CLSI recommendations, phenotypic antibiotic susceptibility testing revealed that strain 022AE was susceptible to all seven tested antibiotics, including chloramphenicol. These results were previously noted for other strains of L. plantarum as well, confirming the strain's safety profile.
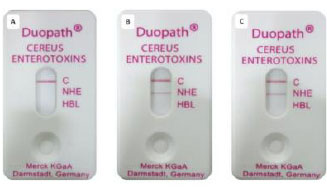
Image showing Duopath® device after 30 minutes of incubation with respective samples. A: Lactobacillus plantarum 022AE; B: Bacillus cereus ATCC 14579; C: Bacillus cereus ATCC11778.
| Treatment | Fluorescence Measurement | % Fluorescence with Respect to Positive Control |
|---|---|---|
| Positive control (0.1% Triton X-100) | 129.29 | 100 |
| Negative control | 4.81 | 3.72 |
| Background | 0.59 | 0.46 |
| 10 µL (test sample) | 21.30 | 16.48 |
| 50 µL (test sample) | 15.55 | 12.03 |
| 100 µL (test sample) | 14.65 | 11.33 |
| Antibiotic Agent |
Streptococcus Pneumoniae ATCC 49619 |
Lactobacillus Plantarum strain 022AE |
||||
|---|---|---|---|---|---|---|
|
MIC range µg /ml (CLSI, 2012b) |
MIC µg/ml |
Interpretation | MIC Breakpoints (µg/ml (EFSA, 2012) | MIC µg/ml | Interpretation | |
| Clindamycin | 0.03-0.12 | 0.06 | S | 2 | 0.015 | S |
| Chloramphenicol | 2-8 | 4 | S | 8 | 1 | S |
| Ampicillin | 0.06-0.25 | 0.25 | S | 2 | 0.03 | S |
| Gentamicin | # | # | # | 16 | 16 | S |
| Tetracycline | 0.06-0.5 | 0.25 | S | 32 | 0.03 | S |
| Kanamycin | # | # | # | 64 | 32 | S |
| Vancomycin | 0.12-0.5 | 0.125 | S | ## | ## | ## |
| Erythromycin | 0.03-0.12 | 0.125 | S | 1 | 1 | S |
| Gene | Function | Remarks |
|---|---|---|
| 347 genes | General antibiotic resistance | Involved in defence, metabolism, transcription, transport, and cell wall biogenesis |
| DUT87_14410 | Chloramphenicol acetyltransferase | Intrinsic resistance, no mobile element |
| - | Tetracycline resistance MFS efflux pump | Intrinsic resistance, no mobile element |
| - | Daunorubicin resistance protein | Intrinsic resistance, no mobile element |
3.7. Virulence Factors And Biogenic Amine
The genome contained 377 potential virulence factor proteins, the majority of which were involved in metabolic activities, cell wall biosynthesis, and signal transduction. Most importantly, no unfavourable virulence genes, such as invasion proteins or toxins, were found. Table 6 provides a detailed categorization of the genome. The majority of discovered genes were associated with extracellular structures and adhesion, both of which are advantageous for probiotic activity. Glutamate decarboxylase was the only decarboxylase gene identified, and it was not linked to harmful biogenic amines (Table 7).
3.8. Adverse Metabolite Genes
Genes encoding enzymes, mentioned in Table 8, were discovered to be potentially hazardous metabolites. However, there were no genes for arylsulfatase or β-glucuronidase. There is no evidence that these metabolic enzymes in L. plantarum have an adverse effect.
3.9. Prophage And Mobile Element Analysis
Based on scoring criteria, PHASTER analysis found three prophage areas in the genome: 2 complete (prophage 2 and 3) and 1 incomplete (prophage 1). The absence of crucial genes needed for full phage assembly and functionality was discovered by thorough annotation of these areas, confirming the defective and non-functional nature of these prophage sequences, as depicted in Table 9.
Using the ISfinder and ACLAME databases, mobile element analysis identified 119 genomic areas, including 25 insertion sites (ISP2, IS153, and ISLmo8), having considerable resemblance to known mobile elements, as depicted in Table 10. Genes producing biogenic amines, pathogenicity, or antibiotic resistance were not found near these mobile elements, indicating that gene mobility may not pose any risks.
| Category | No. of proteins | Function |
|---|---|---|
| Inorganic ion transport and metabolism | 65 | Nutrient transport |
| Defence mechanisms | 60 | Stress response |
| Cell wall/membrane biogenesis | 44 | Structural element |
| Signal transduction | 34 | Cell signalling |
| Post-translational modification | 22 | Protein processing |
| Others | 152 | Various transport mechanisms, metabolism, cell motility, etc. |
| Gene | Function | Safety concern |
|---|---|---|
| DUT87_06365 | Glutamate decarboxylase | No toxic biogenic amine production |
| Enzyme | No. of genes | Gene IDs |
|---|---|---|
| Beta-glucosidase | 9 | DUT87_03505, DUT87_03510, DUT87_04565, DUT87_06775, DUT87_05085, DUT87_06790, DUT87_06795, DUT87_09030, DUT87_12850 |
| Nitroreductase | 4 | DUT87_02785, DUT87_04385, DUT87_07390, DUT87_07440 |
| Azoreductase | 2 | DUT87_07495, DUT87_10960 |
| D-lactate dehydrogenase | 2 | DUT87_10630, DUT87_15500 |
| Prophage | Region (genomic coordinate) | Status | Reason |
|---|---|---|---|
| Prophage 1 | 128-13623 | Incomplete | Lacks essential genes |
| Prophage 2 | 14671-66987 | Defective | Missing replication and lysis genes |
| Prophage 3 | 335640-419620 | Defective | Lacks lysogeny and morphogenesis genes |
| Mobile element | No. of sites | Remarks |
|---|---|---|
| ISP2 | 11 | Transposase encoding |
| IS153 | 1 | IS3 family |
| ISLmo8 | 1 | Transposase |
| Total mobile regions | 119 | No resistance or virulence genes nearby |
3.10. Probiotic Functional Genes
The strain 022AE possessed a number of genes linked to probiotic traits, like adhesion, bile and acid tolerance, and resistance to environmental stress. Adhesion-related genes include several LPXTG-motif anchored cell wall proteins, fibronectin-binding protein (DUT87_14440), and mucus-binding proteins (DUT87_05005, DUT87_13795), as depicted in Table 11. Chaperonins, Clp protease, pyruvate kinase, and ATP synthase subunits are among the genes that contribute to acid tolerance. Genes, such as peptidoglycan endopeptidase, orotidine 5'-phosphate decarboxylase, and dihydrolipoyl dehydrogenase, have been linked to bile tolerance.
Additionally, the genome included genes encoding stress response proteins, like cold shock proteins, GroEL, GroES, and Hsp33, enabling the strain's adaptability to a variety of environmental circumstances. These genetic characteristics demonstrated the strain's functional safety and probiotic potential, as depicted in Table 12.
3.11. Comparative Genomics
There were 16 rRNA genes predicted from the assembled genome. Among the 16 rRNA genes, six were identified as 5S rRNA, five as 16S rRNA, and five as 23S rRNA. The five 16S rRNA sequences were aligned to the SILVA 16S database and showed significant homology. In
| Gene | Function |
|---|---|
| DUT87_05005 | Mucus-binding protein |
| DUT87_13795 | Mucin-binding domain |
| DUT87_14440 | Fibronectin-binding protein |
|
DUT87_08825 DUT87_02365 DUT87_02775 DUT87_04210 DUT87_04270 DUT87_04415 DUT87_04520 DUT87_04765 DUT87_10840 DUT87_10920 |
LPXTG cell wall anchor proteins Cell wall anchor proteins LPXTG cell wall anchor proteins Cell wall anchor proteins Cell wall anchor proteins Cell wall anchor proteins Cell wall anchor proteins LPXTG cell wall anchor proteins Cell wall anchor proteins Cell wall anchor proteins |
| DUT87_09250, DUT87_00650 | Sortase |
| DUT87_10265 | Enolase |
| Gene | Function |
|---|---|
| Acid Tolerance | |
| DUT87_01770 to DUT87_01805 | ATP synthase subunits |
| DUT87_00225 | Clp protease |
| DUT87_14855 | Pyruvate kinase |
| DUT87_03915 | Phosphoglycerate mutase |
| DUT87_09965 | GroEL |
| DUT87_10785 | GMP synthase |
| Bile Tolerance | |
| DUT87_00700 | Bile tolerance |
| DUT87_03170 | Bile tolerance |
| DUT87_08150 | Bile tolerance |
| DUT87_00750 | Bile tolerance |
| Environmental Stress | |
| DUT87_09965 | GroEL |
| DUT87_09960 | GroES |
| DUT87_09420 | Hsp33 |
| DUT87_15365 | GrpE |
| DUT87_12280 | Clp protease |
| DUT87_06800 | Cold shock protein |
addition, the contig of these 5 16S rRNA sequences was prepared, and BLASTN was carried out, which showed 100% identity to Lactobacillus plantarum. A partial 16S rRNA sequence was deposited in the NCBI/GenBank database under the accession number MH532530.
The assembled genome of L. plantarum strain 022AE was compared with other bacterial genomes present in the RefSeq genome database using NCBI-BLASTN. L. plantarum (taxid: 1590) was chosen as the reference database for NCBI-BLASTN. The BLASTN results indicated 100% sequence homology of the de-novo assembled genome with the genome of the reference strain L. plantarum HAC01. Fig. (6) shows a BRIG circular plot constructed using two genomes, L. plantarum strain 022AE and L. plantarum strain HAC01, using BRIG version 0.95.
4. DISCUSSION
The beneficial effects of probiotic bacteria are strain-specific. Therefore, each probiotic strain must be evaluated at genetic, phenotypic, pre-clinical, and clinical levels. The probiotic potential of numerous bacterial and fungal strains has been discovered with WGS technology. The identification of genes that contribute to probiotic characteristics and those that may raise safety concerns is made easier by the thorough annotation of assembled genomes. In this study, we used genetics and in vitro studies to evaluate the safety and probiotic potential of L. plantarum 022AE. 16S rRNA gene sequencing was used to identify L. plantarum 022AE. L. plantarum 022AE entire genome was sequenced with a coverage of more than 200 times. The genome size of L. plantarum was a single scaffold of 3,234,271 bp with the GC content of about 44.55%. Both the genome size and GC content fell in the range reported for this species by Altschul, Pruesse, and Ali Khan [28-30].
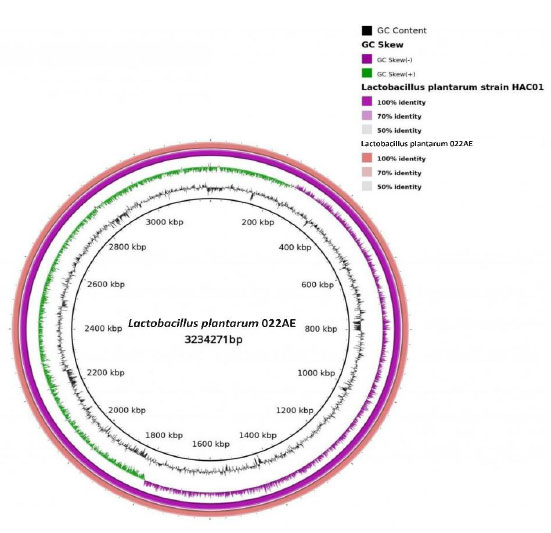
Circos plot comparison of L. plantarum strain 022AE (size of genome: 3,234,271 bp) with L. plantarum strain HAC01 (CP029349.1).
Genetic rearrangements and horizontal gene transfer can influence bacterial genomes on an evolutionary time scale. Antibiotic-resistance genes or virulence factors are captured, accumulated, and disseminated by mobile elements, such as insertion sequences. None of the insertion sequences was found near the predicted virulence factors or antibiotic resistance genes in the 022AE assembled genome. Prophage sequence analysis revealed the presence of three prophage regions, but they were annotated as defective and non-functional. Approximately 40% of sequenced bacterial genomes contain CRISPR short direct repeats (23-47 bp in length) that help regulate genome stability by offering immunity against plasmids and bacteriophages encountered in the past. Each of these repeats is separated by spacers of similar length that are specific to each genome and show the non-coding section of genomic sequences between the genes. The L. plantarum strain 022AE genome has four CRISPR and one Cas gene, which suggests an advantage in ensuring genome stability by preventing the entry of foreign DNA elements [5].
After oral intake, probiotics encounter acidic gastric juice, bile salts, and digestive enzymes. Hence, survival in these environments is a fundamental probiotic attribute. L. plantarum 022AE showed survival at pH 3.5 for 5 hours and tolerated bile concentrations up to 1.0%. It also survived through simulated gastrointestinal (GI) conditions, both in fed and fasting models. Diets, like SAD and SED, improved its survival. The identification of genes associated with survival under these stresses further confirmed the strain’s robustness in such environments (Table 12). The strain L. plantarum GXL94 and others showed similar bile and acid tolerance, i.e., 96% viability at pH 2.5 and 95% at 1.0% bile, as described by Yingjun. L. plantarum E680 strain was studied by Zhi-Yao in 2020, and strains B2, YJ24, YJ14, and HN9 were studied by Hao in 2021 [31-34]. However, such tolerance depends on strain origin and conditions.
Acid and bile stress resistance in L. plantarum involves proteins, like GrpE, MetE, RpsB, ClpP, Dps, GroEL, Hsp1, Hsp3, GshR1, GshR4, and OpuA, as described by E. Hamon [35, 36]. Furthermore, Hamon et al. discovered proteins of L. plantarum in bile salt response, which included glutathione oxidases, fatty acid phospholipid synthase, bile salt hydrolase, and ATP synthase [35, 36]. The L. plantarum 022AE-assembled genome also contained ATP synthase subunits and amino acid decarboxylase, which may aid in the regulation of bile-related stress factors by preserving H+ homeostasis and alkalinizing the cytosol, thus affecting its ability to endure in the acidic gut environment [31]. Lactobacillus plantarum 022AE exhibited beneficial cell surface properties, including autoaggregation (the accumulation and precipitation of cells to prevent rapid removal during intestinal peristalsis), co-aggregation with key pathogens, and adhesion to nonpolar solvents, indicating its capacity to attach to the hydrophobic surfaces of gut epithelial cells. This has been found to be consistent with studies on L. plantarum BBC33, which have shown 24.8% xylene affinity and 37.2% autoaggregation, as explained by Bharat [37].
Additionally, L. plantarum 022AE produced β-galactosidase, which may help reduce lactose intolerance, and bile salt hydrolase (BSH) that may lower cholesterol [38]. The authors, like Anna, Nirunya, C.G. Vinderola, and JH Miller, have explained similar properties [39-42]. The strain showed antioxidant activity in cell-free extracts by scavenging DPPH free radicals, indicating its potential as a postbiotic. Agolino Gianluigi et al. reported similar observations for L. plantarum DMDL 9010 [43]. Namrata Bhingardeve demonstrated the antimicrobial activity of the L. plantarum 022AE strain. Its cell-free extracts inhibited pathogens, like Bacillus subtilis ATCC 6633, C. perfringens ATCC 13124, B. cereus ATCC 33019, and Listeria monocytogenes ATCC 19115 [3]. Jones Mitchel reported similar anti-listerial activity of L. plantarum Q7, and Echegaray Noemi reviewed several bacteriocins from L. plantarum. Genome analysis of L. plantarum 022AE confirmed the presence of plantaricin genes [38, 44]. The strain showed acceptable industrial potential. It resisted heat stress with a D-value of 1.54 h at 50°C and maintained 95% viability between 4-40°C for 6 hours, as explained by Namrata Bhingardeve [3]. Similar results were obtained with other strains of L. plantarum described by Liu Dong Mei and Liu Yinxue [45, 46]. It also remained stable in buffer solutions for up to 6 months under refrigerated (97.8% viability) and ambient (89.5% viability) conditions. Overall, L. plantarum 022AE showed excellent in vitro probiotic features, including GI tolerance, adhesion, antimicrobial and antioxidant activity, enzyme production, and industrial stability. These findings suggest its potential for human and animal nutrition.
Adhesion to the intestinal mucosa is crucial for colonization and has been deemed an ideal parameter for determining a prospective probiotic strain's harbouring capabilities. Attachment of probiotic bacteria to the gastrointestinal surface increases their residency length in vivo, influencing host health by boosting the gut immune system; it may also be a preliminary step for competitive exclusion of harmful bacteria [14]. The presence of mucin adherence in the 022AE strain suggested the possibility of gut colonization. Several previous studies have used human epithelial cell lines, such as Caco-2, to assess the adhesion properties of probiotic strains [14]. These cellular models have the morphological and functional properties of mature enterocytes and express the majority of the receptors, enzymes, and transporter proteins found in the normal human intestinal epithelium [47]. According to recent studies, L. plantarum Lp91 showed 10.2%, L. plantarum CIAL 121 showed 7.10%, L. plantarum AdF10 showed 9.55%, L. plantarum AdF5 showed 8.33%, and L. plantarum AdF6 showed 7.37% adherence to Caco-2 cell lines [2, 13, 14]. A study by Echegaray Naomi reported a strain with the highest adhesive potential, i.e., L. plantarum WEH 92 [44]. Its cell wall showed overexpression of proteins, like protein elongation factor, GroEL chaperonin, GroES co-chaperonin, and glyceraldehyde-3-phosphate-dehydrogenase, and hence, linked with adhesion ability. Maurits van den Nieuwboer described similar adhesion-specific genes in L. plantarum WCFS1 [31, 48]. L. plantarum 022AE also showed the presence of GroEL and GroES (DUT87_09965 and DUT87_09960) and other gene clusters that facilitate adhesion to gut mucosa, as mentioned in Table 11. These genes, together with the adhesion of Lactobacillus plantarum 022AE to mucin and Caco-2 cell lines in in vitro assays, indicate its potential for efficient colonization and survival in the gastrointestinal tract.
In an in vitro test for assessing the cytotoxicity against Vero cells, culture supernatants of L. plantarum 022AE showed no cytotoxic effect. The metabolites of the majority of Lactobacillus strains exhibited little to no cytotoxicity towards Caco-2 cells, according to Katarzyna Slizewska [17]. The absence of enterotoxins in culture supernatants of L. plantarum 022AE and its susceptibility to CLSI-recommended antibiotics further strengthen its safety assessments. According to Yoo Jin Kwon, L. plantarum strains Q180 and DSM 20174 did not produce enterotoxins [49].
CONCLUSION
Lactiplantibacillus plantarum strain 022AE has a larger genome than those of the other LAB species. This reflects the ability of the species to inhabit a wide range of environments, which is further associated with strain-specific variations observed in several attributes of this bacterium. This scenario necessitates a thorough examination of the strains of this species across as many habitats as possible to better understand the intriguing and diverse phenotypes it exhibits. To our knowledge, this is the first study to combine genomic and phenotypic characteristics of the L. plantarum strain. Further research is being conducted to functionally characterize biotechnologically relevant genes in these isolates, shedding light on yet-unknown aspects of L. plantarum. The findings from a comparative genomic analysis and phenotypic attributes highlight the importance of establishing genotype-phenotype correlations for a broader range of traits to better understand and utilize this fascinating bacterium.
AUTHORS' CONTRIBUTIONS
D.S. was responsible for conceptualizing the project and reviewing the paper. A.I. examined the manuscript and made revisions. P.U. performed the experiment involving WGS. G.C., P.U., and A.C. compiled the data. A.C. wrote the manuscript's initial draft and did the editing. All the authors reviewed the results and approved the final version of the manuscript.
LIST OF ABBREVIATIONS
| LAB | = Lactic acid bacteria |
| GRAS | = Generally recognized as safe |
| QPS | = Qualified presumption of safety |
| WGS | = Whole-genome sequencing |
| CARD | = Comprehensive Antibiotic Resistance Database |
AVAILABILITY OF DATA AND MATERIALS
The material generated during and/or analyzed during the current study will be available from the corresponding author upon reasonable request.
FUNDING
This work was supported by the Research and Development Department of Advanced Enzymes Technologies Ltd., India.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge the support from Mr. V.L. Rathi and Mr. M. Kabra at Advanced Enzyme Technologies Ltd., India.

