All published articles of this journal are available on ScienceDirect.

Description of the Cariogenic and Periodontopathogenic Microbiome in Patients with and Without Oral Squamous Cell Carcinoma Using Next-Generation Sequencing
Authors Info & Affiliations
Abstract
Introduction
Periodontal disease and dental caries are chronic conditions that share a common risk factor: the bacterial biofilm, which is associated with inflammation and may also be linked to malignant transformation. This study determined the composition and qualitative and quantitative differences in the cariogenic and periodontopathogenic microbiomes of patients with and without oral squamous cell carcinoma (OSCC).
Methods
A case-control study was conducted using plaque and saliva samples from 10 OSCC patients and 10 healthy controls. DNA was extracted and sequenced using the Illumina MiSeq platform. Bioinformatics tools and reference databases were employed for taxonomic classification. Bacterial identification was based on a predefined profile of 140 periodontopathogenic and 111 cariogenic species. Analyses included intergroup comparisons using the Mann-Whitney U test, principal coordinate analysis, and descriptive statistics (means, standard deviations, and frequencies) to assess differences between groups.
Results
Within the defined cariogenic and periodontopathogenic microbiome profiles, 26 and 20 bacterial species, respectively, were identified in the total plaque and saliva samples from both study groups. A significant number of the 46 identified bacteria were shared by both groups with a high frequency and abundance. Lactobacillus helveticus, L. reuteri, Streptococcus pasteurianus, S. parauberis, S. infantarius, S. lutetiensis, S. macedonicus, and Enterobacter asburiae were found to be differentially abundant.
Discussion
Some bacterial species may play a potential eubiotic role, and some were found at low frequency in the saliva of patients and at high frequency in the saliva of controls.
Conclusion
A cariogenic and periodontopathogenic microbiome profile was established, and eight bacteria with potential eubiotic impact are highlighted.
1. INTRODUCTION
It is estimated that oral biodiversity comprises over 700 bacterial species that coexist in complex communities in the form of biofilms. These are typically composed of microbial species embedded in an extracellular matrix, along with proteins, polysaccharides, lipids, salivary glycoproteins, albumin, and host cellular components [1]. The homeostasis in which bacteria coexist within different ecological niches can be disrupted by various host-related factors, potentially leading to an ecological imbalance in the bacterial community, which in turn may result in oral disease [2]. Several types of diseases can largely result from bacterial dysbiosis, as the number of symbionts beneficial to normal tissue function decreases while pathogenic species increase [3]. This imbalance may promote the release of inflammatory mediators and lead to mutagenesis, uncontrolled cell proliferation, angiogenesis, epigenetic modifications, and, in some cases, cancer [4]. Poor oral hygiene has been clearly established as a risk factor for oral cancer, as it leads to a qualitative and quantitative increase in microorganisms [2]. Some studies have reported that oral hygiene status influences not only the onset but also the progression and promotion of oral, oropharyngeal, and laryngeal cancers, among others [5]. Possible explanations include the observation that chronic inflammatory processes may create hostile microenvironments for cells, increasing cell proliferation, mutagenesis, oncogene activation, and angiogenesis [6]. Given this context, it is essential to study the composition of bacterial communities and to describe, both qualitatively and quantitatively, the microorganisms present in different oral niches. Some of these bacteria may serve as potential biomarkers for clinical and microbiological diagnosis in health and disease [7]. Periodontal disease and dental caries are chronic infectious pathologies with different clinical manifestations. Periodontal disease is characterized by proinflammatory symptoms that alter the supporting structures of the teeth, while dental caries is characterized by the demineralization and destruction of enamel and dentin [8, 9]. It is very important to note that these two pathologies have as a risk factor the formation of a bacterial biofilm made up of multiple microorganisms from the oral microbiome [10, 11]. A healthy oral microbiome, or one compatible with eubiosis, plays a crucial role in maintaining systemic health. It modulates immune responses, influences nutrient metabolism and appetite, contributes to vitamin biosynthesis, protects against exogenous pathogens, and produces antimicrobial agents [12]. When this balance is disrupted, dysbiosis occurs, favoring pathogenic bacteria that can contribute to diseases such as gingivitis, periodontitis, dental caries, and potentially oral cancer [12-14]. The composition and abundance of the microbiome in the oral cavity differ from one microenvironment to another [15]. The onset of periodontal disease is characterized by an increase in pathobionts and key pathogens such as Fusobacterium nucleatum, Porphyromonas gingivalis, Prevotella intermedia, and Aggregatibacter actinomycetemcomitans, among other microorganisms, which can alter the host immune response and contribute to the development of periodontal disease [16, 17]. On the other hand, in dental caries, bacteria with acidophilic and aciduric properties play an important role in the initiation and progression of the carious lesion. Bacteria like Streptococcus mutans, Streptococcus salivarius, Streptococcus sanguinis, and species of Actinomyces and Lactobacillus are the most frequently associated with dental caries and can vary qualitatively and quantitatively depending on the stage of the carious lesion [10]. In this sense, it is of utmost importance to control the formation and accumulation of subgingival and supragingival biofilm to maintain symbiosis and improve the balance of oral biofilms [16]. If these pathogens are present in the biofilms, they will induce chronic inflammatory responses that increase the release of cytokines and inflammatory mediators and ultimately alter the dental tissues [18-20]. It has been shown that persistent inflammation has a strong association with the malignant transformation of the affected oral epithelium and that the presence of bacteria, such as Fusobacterium periodonticum, F. nucleatum, P. intermedia, Aggregatibacter segnis, Pseudomonas aeruginosa, Peptostreptococcus stomatis, Tannerella forsythia, Catonella morbi, and A. actinomycetemcomitans, could play an important role in the development of oral squamous cell carcinoma (OSCC) [14, 21]. Studies such as that of Wolf et al. in 2017 [22], in which a comparative analysis of the microbiome was performed in saliva samples from 11 patients with OSCC and 11 healthy patients using high-throughput sequencing of the 16S rRNA gene, allowed researchers to find differences in the salivary microbiome between these two groups of patients. The genera Prevotella, Streptococcus, Haemophilus, Veillonella, and Neisseria were found with the highest abundance in healthy controls, while Actinomyces, Schwartzia, Treponema, and Selenomonas were the most abundant in patients with OSCC. These qualitative and quantitative differences may reflect the importance of some bacteria in disease progression. In another investigation, Dong et al. (2018) [23] described the set of microbial communities present in different sites of the oral cavity and esophagus, sequencing the V3-V4 region of the 16S rRNA gene in clinical samples of saliva, the dorsum of the tongue, supragingival plaque, and upper, middle, and lower segments of the esophagus in 27 healthy individuals, and found that the genera Actinobacillus, Prevotella, and Veillonella were the most abundant (>1%) in the oral cavity and esophagus. In contrast, the genera Neisseria and Streptococcus were found, respectively, with greater frequency in the oral cavity and esophagus. Guerrero-Preston et al. (2016) [24] conducted a study using 16S rRNA V3-V5 gene sequencing to compare the saliva microbiome from patients with OSCC and normal epithelium controls before and after surgical resection to characterize and examine their abundance. The researchers frequently found the phyla Firmicutes, Proteobacteria, and Bacteroidetes, and less frequently Actinobacteria and Fusobacteria before surgery. On the other hand, it was found that microbial alpha diversity decreased after surgery, but this diversity increased in patients with recurrence (p<0.05). However, despite the various studies carried out, there is no consensus on the interpretation of these findings and their clinical use. Deeper sequencing studies with enhanced resolution and analysis are required to elucidate the precise composition and interactions within the OSCC microbial community at the species level [25-27]. A comprehensive understanding of microbiome dynamics across health and disease states is essential to identify microorganisms differentially associated with OSCC and to explore their potential as biomarkers for prevention and control strategies targeting oral diseases, including OSCC [21, 26]. Given this situation, the objective of this study was to describe the composition and qualitative and quantitative differences in the cariogenic and periodontopathogenic microbiomes present in patients with and without OSCC. Microbiome analysis can be approached through 16S rRNA gene sequencing and shotgun metagenomics. The 16S rRNA-based technique allows for the identification and classification of bacteria primarily at the genus level, whereas shotgun metagenomics sequences all genetic material present in the sample, leading to a deeper and more accurate characterization of the microbial community. Metagenomics enables the identification of the taxonomic composition and functional potential of microbial communities. Meanwhile, metatranscriptomics analyzes messenger RNA, providing information on the genes that are actively expressed at a given time. Both techniques are complementary, and their integration offers a more comprehensive view of the microbial ecosystem, which is key to understanding its functional dynamics and response to environmental or clinical factors. Therefore, it is necessary to advance the implementation of new technological tools that integrate metagenomic and metatranscriptomic studies in order to more precisely describe the composition, interaction, and activity of the bacterial community in different oral microenvironments of patients with and without OSCC [28, 29]. Therefore, the objective of this study was to describe the composition and the qualitative and quantitative differences in the cariogenic and periodontopathogenic microbiomes of patients with and without OSCC.
2. MATERIALS AND METHODS
A case-control study was conducted using saliva, dental plaque, and tumor samples from 10 patients diagnosed with oral squamous cell carcinoma and saliva and dental plaque samples from 10 control individuals without OSCC. This study was conducted in accordance with the ethical principles outlined in the Declaration of Helsinki, after obtaining approval from the bioethics committee and informed consent from all participants [CIEFOPUJ Act 11 of 2015]. Patients of both sexes and in any age group were included, provided they had a confirmed diagnosis of OSCC established through histopathological analysis independently reviewed by two expert pathologists and had at least four natural teeth. Control group participants were selected by matching them for age and sex with the patient group and had no oral pathologies or history of orthodontic treatment. Exclusion criteria included the use of oral antiseptics (e.g., mouthwashes) within two days prior to sample collection, antibiotic use within two months prior, and poor-quality DNA sequences. All samples were collected following the guidelines established in the protocols described for studies of the Human Microbiome Project [30] and were collected by a single, previously calibrated operator. Each patient deposited approximately 5 mL of unstimulated saliva into a tube containing RNAlater buffer to preserve nucleic acids. An odontogram was performed for all participants to record the presence or absence of teeth. Additionally, without brushing the night before, the modified Silness and Löe Index was applied to assess the level of dental plaque. Subsequently, a supragingival plaque sample was collected using a scraping technique with Gracey Micro Mini Five curettes, with field isolation using cotton rolls and drying with a triple-syringe air jet. The dental surfaces evaluated included the buccal, distal, palatal/lingual, and distal surfaces of teeth 16, 21, 24, 36, 41, and 44. In cases where any of these teeth were missing, the sample was taken from the mesial surface of the adjacent tooth. In patients with only four teeth, these were considered index teeth. Plaque samples were collected in RNAlater buffer. Finally, the tubes were stored at −70 °C until processing.
2.1. DNA Extraction, Quantification, And Quality Analysis
DNA was isolated from all samples using the MasterPure™ DNA Purification Kit (Epicentre Biotechnologies®, Madison, WI), following the manufacturer’s instructions. This method enables high-quality DNA extraction through filtration and enzymatic technology. DNA quantification was subsequently performed by fluorometry using the QuantIT™ dsDNA BR Assay Kit (Invitrogen, Carlsbad, CA, USA), and DNA integrity was assessed via 1% agarose gel electrophoresis. Quality control criteria included a DNA yield greater than 50 ng/μL and a purity ratio (A260/A280) between 1.8 and 2.0. Samples meeting these criteria were submitted for next-generation sequencing using the Illumina MiSeq™ 2500 platform, generating 250 bp paired-end reads.
2.2. Library Preparation, Sequencing, and Quality Control
Genomic libraries were prepared using the Illumina Nextera XT DNA Library Preparation Kit (Illumina Inc., San Diego, CA, USA), following the manufacturer’s protocol. All libraries were quantified and pooled in equimolar concentrations. Automated cluster generation and paired-end sequencing were performed on the Illumina MiSeq™ 2500 platform, according to the manufacturer’s instructions. Library quality was assessed using the Agilent TapeStation D1000 ScreenTape system (Agilent Technologies, Santa Clara, CA, USA). Sequencing generated 250 bp paired-end reads. Raw FASTQ files were processed using Trimmomatic [31] for initial quality filtering. Sequences with a Phred score <20 and reads shorter than 100 bp from the 3′ end were discarded. Nextera adapter sequences were removed, and low-quality nucleotides were trimmed. Further quality control included adapter trimming using Cutadapt (minimum overlap: 3 bp; error rate: 0.1), removal of chimeric sequences using the de novo chimera detection algorithm in DADA2, and quality assessment and visualization using FastQC (available at: https://www.bioinformatics.babra ham.ac.uk/projects/fastqc/). Taxonomic profiling of bacterial communities was performed using Kraken v1.0 [32] with the Kraken DB database, which includes complete bacterial genomes from RefSeq [available at: https://ccb.jhu.edu/software/kraken/MANUAL.html#kraken-databases]. Kraken was selected for its computational efficiency and accuracy in handling diverse oral microbiomes compared to alignment-based methods used with databases like SILVA or Greengenes. The Human Oral Microbiome Database (HOMD) was used as the reference due to its comprehensive coverage of oral bacterial taxa, with a confidence threshold of 0.8 for taxonomic assignments.
2.3. Statistical Analysis
To account for rare or low-abundance taxa, sequence data were normalized by rarefaction to a uniform sequencing depth of 10,000 reads per sample. Intergroup comparisons of bacterial abundance were performed using the Mann-Whitney U test, with p-values adjusted for multiple testing using the False Discovery Rate (FDR) correction (Benjamini-Hochberg method). DNA sequence data were processed into a feature table of microbiome composition, with bacterial taxa indexed by rows and metagenomic sample abundances tabulated across columns. A principal coordinate analysis (PCoA) was performed using Bray-Curtis dissimilarity on a transposed table with metagenome samples as rows and bacterial counts as columns, derived from the original feature table of microbiome composition.
Finally, the information was analyzed using averages, standard deviations, and frequencies of bacteria present in the samples studied to determine if there was a relationship between the presence of cariogenic and periodontopathogenic bacteria in the saliva and plaque samples of patients with and without OSCC. To identify bacteria associated with dental caries and periodontal disease, a database search and literature review of studies involving plaque and saliva sequencing from patients with these diseases were conducted. Subsequently, these bacteria were searched within the metagenome database of plaque and saliva samples from patients with and without OSCC.
3. RESULTS
3.1. Characteristics of Patients With And Without OSCC
The study sample was predominantly female (90%), with a mean age of 56 years. Significant differences were evident in oral hygiene habits: 80% of the control group reported good oral hygiene, while only 20% of the patients indicated the same. Conversely, 60% of the patients exhibited fair and poor oral hygiene. Concerning tumor location, 50% were on the tongue, followed by 30% in the cheek and 20% on the lips (Table 1).
3.2. Cariogenic Bacterial Profile In Plaque And Saliva Samples Of Patients With And Without OSCC
To establish cariogenic bacterial profiles present in dental plaque and saliva samples obtained from patients with and without OSCC, a comprehensive literature search was conducted focusing on previous studies that employed DNA sequencing of plaque and saliva samples from patients with and without caries. A total of 111 bacterial species were identified within the cariogenic profile (Supplementary Appendix 1). The plaque and saliva samples of patients with and without OSCC were screened for the presence of these bacterial species. A total of 26 bacterial species were identified, including: Atopobium parvulum, Lactobacillus gasseri, Lactobacillus helveticus, Lactobacillus reuteri, Lactobacillus salivarius, Leptotrichia buccalis, Rothia dentocariosa, Rothia mucilaginosa, Selenomonas sputigena, Streptococcus anginosus, Streptococcus constellatus, Streptococcus cristatus, Streptococcus gordonii, Streptococcus infantarius, Streptococcus intermedius, Streptococcus lutetiensis, Streptococcus macedonicus, Streptococcus mitis, Streptococcus mutans, Streptococcus oralis, Streptococcus parasanguinis, Streptococcus parauberis, Streptococcus pasteurianus, Streptococcus salivarius, Streptococcus sanguinis, and Veillonella parvula.
The cariogenic bacterial profile in patients with and without OSCC is depicted in Fig. (1). The 26 bacterial species exhibited varying prevalence rates across the two study groups. In OSCC patients (Fig. 1A), V. párvula, S. sanguinis, S. salivarius, S. parasanguinis, S. oralis, S. mitis, S. intermedius, S. gordonii, S. cristatus, S. constellatus, S. anginosus, S. sputigena, L. buccalis, A. parvulum, and R. dentocariosa were highly prevalent (>80%) in both plaque and saliva. Less frequent species included R. mucilaginasa and L. salivarius (60% in saliva vs. 100% in plaque), S. mutans (60% in saliva vs. 90% in plaque), L. gasseri (60% in both sample types); S. macedonicus (50% in saliva vs. 80% in plaque), S. lutetiensis (40% in saliva vs. 80% in plaque), Lactobacillus helveticus (10% in saliva vs. 80% in plaque), L. reuteri (20% in saliva vs. 70% in plaque), S. parauberis (30% in saliva vs. 60% in plaque), S. infantarius (30% in saliva vs. 50% in plaque), and S. pasteurianus (30% in both sample types).
| Group | No. | Age years | Sex | Smoke | Alcohol intake | Tumor localization | Oral health |
|---|---|---|---|---|---|---|---|
| OSCC group | 10 | Mean 56.4 *SD ± 18.36 |
F = 9 M = 1 |
Yes = 1 No = 9 |
Yes =1 No = 9 |
Tongue = 5 Cheek = 3 Lip = 2 |
Good = 2 Regular = 6 Bad = 2 |
| Control group | 10 | Mean 56.5 *SD ± 15.59 |
F = 9 M = 1 |
Yes = 1 No = 9 |
Yes = 1 No = 9 |
NA | Good = 8 Regular = 2 Bad = 0 |
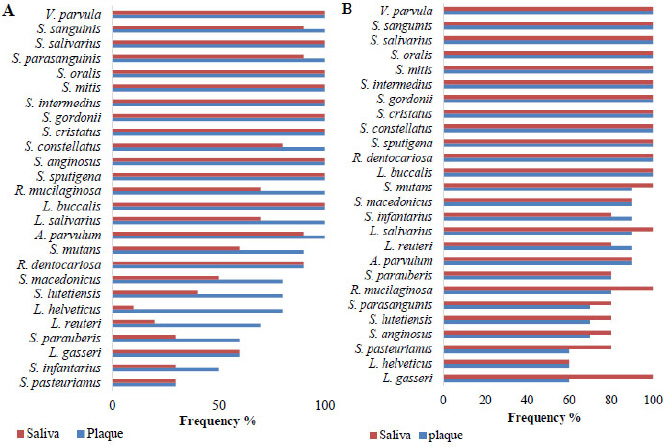
Cariogenic bacterial profile. A. Frequency in the plaque and saliva of patients with OSCC. B. Frequency in the plaque and saliva of patients without OSCC.
The cariogenic profile in patients without OSCC is depicted in Fig. (1B). The following species exhibited high prevalence rates (>80%) in both plaque and saliva samples: V. parvula, S. sanguinis, S. salivarius, S. oralis, S. mitis, S. intermedius, S. gordonii, S. cristatus, S. constellatus, S. sputigena, R. dentocariosa, L. buccalis, S. mutans, S. macedonicus, S. infantarius, L. salivarius, L. reuteri, A. parvulum, S. parauberis, and R. mucilaginosa. Less frequent species included S. parasanguinis, S. lutetiensis, and S. anginosus (70% in plaque vs. 80% in saliva), S. pasteurianus (60% in plaque vs. 80% in saliva), L. helveticus (60% in plaque vs. 60% in saliva), and L. gasseri (60% in plaque vs. 100% in saliva).
A differential analysis was conducted to identify bacterial species with potential eubiotic properties. Specifically, we sought bacterial species that were less prevalent in the saliva of patients with OSCC compared to those without OSCC. The following species were identified: L. helveticus (10% versus 60%), L. reuteri (20% versus 80%), S. pasteurianus (30% versus 80%), S. parauberis (30% versus 80%), S. infantarius (30% versus 80%), S. lutetiensis (40% versus 80%), and S. macedonicus (50% versus 90%) (Fig. 2).
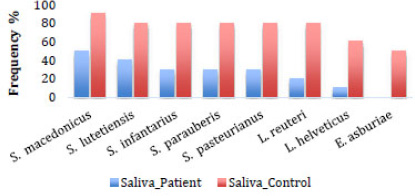
Bacterial profile associated with eubiosis. Frequency of bacterial species in the saliva samples of patients with and without OSCC.
3.3. Periodontal Pathogenic Bacterial Profile In Plaque And Saliva Of Patients With And Without OSCC
To characterize the periodontopathogenic bacterial profiles in dental plaque and saliva samples from patients with and without OSCC, a comprehensive literature review was conducted, focusing on previous studies employing DNA sequencing of samples from patients with periodontal disease. A total of 140 bacterial species were identified within the periodontopathogenic profile (Supplementary Appendix 2). A comprehensive search was conducted for these bacterial species in plaque and saliva samples obtained from patients with and without OSCC. A total of 20 bacterial species were identified, namely Capnocytophaga canimorsus, Gardnerella vaginalis, Providencia stuartii, Enterobacter asburiae, Campylobacter concisus, Klebsiella pneumoniae, Pseudomonas aeruginosa, Serratia marcenses, Filifactor alocis, Treponema denticola, Aggregatibacter actinomycetemcomitans, Aggregatibacter aphrophilus, Capnocytophaga ochracea, Fusobacterium nucleatum, Haemophilus parainfluenzae, Porphyromonas gingivalis, Prevotella intermedia, Prevotella melaninogenica, Tannerella forsythia, and Prevotella denticola.
Fig. (3) presents the periodontopathogenic bacterial profile found in patients with and without OSCC. The 20 bacterial species were detected at different frequencies in the two study groups. In patients with OSCC (Fig. 3A), the following bacteria were found with high frequency (>80%) in both types of samples: P. denticola, S. marcescens, C. canimorsus, T. forsythia, P. melaninogenica, P. intermedia, P. gingivalis, H. parainfluenzae, F. nucleatum, F. alocis, C. ochracea, A. aphrophilus, and A. actinomycetemcomitans. Less frequently, the following bacteria were observed: K. pneumoniae and T. denticola (both with 60% frequency in saliva versus 90% in plaque), P. aeruginosa (40% frequency in saliva versus 90% in plaque), C. concisus (60% frequency in saliva versus 90% in plaque), P. stuartii (30% frequency in saliva versus 80% in plaque), G. vaginalis (70% frequency in saliva versus 80% in plaque), and E. asburiae (0% frequency in saliva and 60% in plaque).
In patients without OSCC (Fig. 3B), the following species were found with high frequency (>80%) in both types of samples: P. denticola, T. forsythia, P. melaninogenica, P. intermedia, P. gingivalis, H. parainfluenzae, F. nucleatum, C. ochracea, A. aphrophilus, and A. actinomycetemcomitans. Less frequently, the following bacteria were found: T. denticola and F. alocis (both 70% in saliva versus 90% in plaque), S. marcescens (70% in saliva versus 80% in plaque), P. aeruginosa (40% in saliva versus 80% in plaque), K. pneumoniae and C. concisus (both 40% in saliva versus 60% in plaque), E. asburiae (50% in both types of samples), and P. stuartii, G. vaginalis, and C. canimorsus, with frequencies of 20%, 30%, and 40%, respectively, only in saliva samples. As a potential eubiotic, only Enterobacter asburiae was found within the periodontopathogenic profile in saliva samples (0% in patients with OSCC and 50% in patients without OSCC) (Fig. 2).
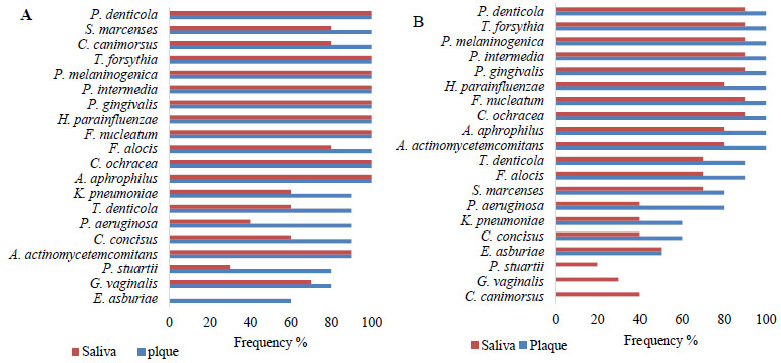
Periodontopathic bacterial profile. A. Frequency in the plaque and saliva of the OSCC group. B. Frequency in the plaque and saliva of the control group.
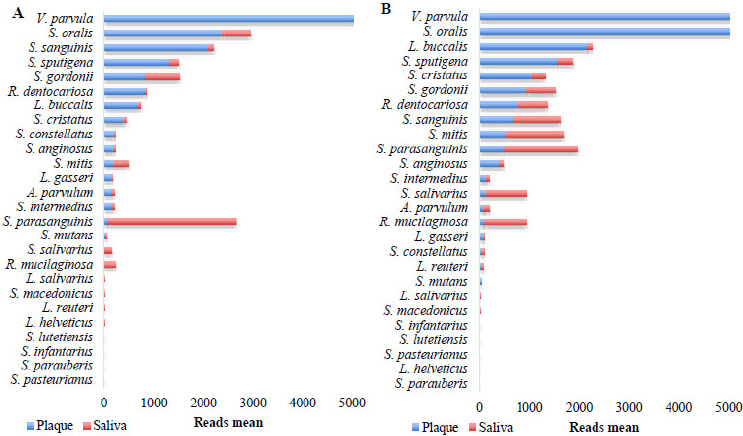
Cariogenic profile. A. Plaque and saliva of the OSCC group. B. Plaque and saliva of the control group.
3.4. Abundance Of Cariogenic Bacterial Species In Dental Plaque And Saliva Of Patients With And Without OSCC
Read abundance analysis revealed distinct bacterial profiles between plaque and saliva in both OSCC and control groups. In OSCC patients, the species V. parvula, S. oralis, S. sanguinis, S. sputigena, R. dentocariosa, L. buccalis, S. cristatus, S. constellatus, S. anginosus, L. gasseri, A. parvulum, and S. intermedius exhibited significantly higher abundance (>150 reads) in plaque compared to saliva (Fig. 4A). Conversely, S. parasanguinis, S. salivarius, and R. mucilaginosa were more prevalent in saliva. In the control group, V. parvula, S. oralis, L. buccalis, S. sputigena, S. cristatus, and R. dentocariosa dominated plaque, while S. sanguinis, S. mitis, S. parasanguinis, S. salivarius, and R. mucilaginosa were enriched in saliva (Fig. 4B). It is noteworthy that in both groups, the species S. mutans did not stand out in terms of abundance (on average 49.8 copies in plaque of patients with OSCC versus 36.89 copies in plaque of patients without OSCC, and 2.8 copies in saliva of patients with OSCC versus 12.2 copies in saliva of patients without OSCC).
3.5. Abundance of Periodontopathogenic Bacterial Species In Dental Plaque And Saliva Of Patients With And Without OSCC
Analysis of periodontopathogenic bacterial profiles revealed distinct distribution patterns between plaque and saliva in both OSCC and control groups. In OSCC patients, P. melaninogenica, C. ochracea, F. nucleatum, P. denticola, A. aphrophilus, P. intermedia, P. gingivalis, T. forsythia, and T. denticola were significantly more abundant in plaque compared to saliva (Fig. 5A). Conversely, H. parainfluenzae was enriched in saliva. In the control group, P. melaninogenica, P. intermedia, C. ochracea, F. nucleatum, T. forsythia, P. gingivalis, and A. aphrophilus predominated in plaque, while H. parainfluenzae, P. denticola, and T. denticola were more prevalent in saliva (Fig. 5B).
3.6. Principal Coordinate Analysis (PCoA)
Beta diversity, evaluated via Bray-Curtis dissimilarity and visualized with principal coordinate analysis (PCoA), revealed distinct microbial community structures (PERMANOVA, p = 0.03), as shown in Fig. (6). Within the cariogenic and periodontopathogenic microbiome profile, 26 and 20 bacterial species were identified, respectively, in plaque and saliva samples from both study groups. Most of the 46 identified species were shared with high frequency and abundance between OSCC and non-OSCC groups. Eight species (Lactobacillus helveticus, L. reuteri, Streptococcus pasteurianus, S. parauberis, S. infantarius, S. lutetiensis, S. macedonicus, Enterobacter asburiae) were differentially abundant, with significantly lower abundance in OSCC patients’ saliva compared to controls (FDR-adjusted p < 0.05, Mann-Whitney U test). Effect sizes (Cohen’s d) ranged from 0.8 to 1.2, indicating moderate to large differences, with 95% confidence intervals reported for each species (e.g., L. helveticus: CI (0.05-0.15)).
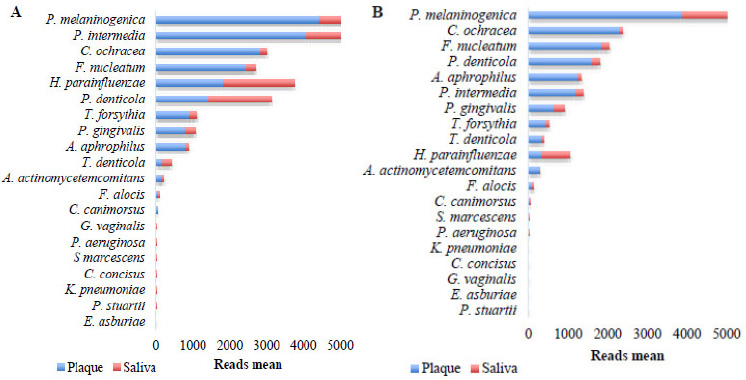
Abundance of periodontopathogenic species. A Plaque and saliva of the OSCC group. B. Plaque and saliva of the control group.
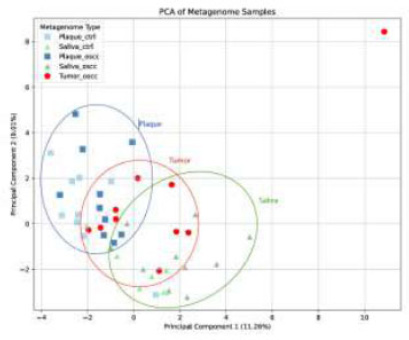
Principal Coordinate Analysis (PCoA). metagenome samples based on Bray-Curtis dissimilarity, showing separation by sample type. Percentages indicate variance explained by principal components 1 (11.26%) and 2 (9.1%).
4. DISCUSSION
Oral squamous cell carcinoma is a globally significant malignancy associated with substantial morbidity and mortality [33]. Oral dysbiosis, characterized by an imbalance in the oral microbiome with a predominance of pathogenic bacteria, has been implicated in the pathogenesis of several oral diseases, including gingivitis, periodontitis, dental caries, and oral cancer [10-12].
Moreover, it is well-established that chronic inflammation is a significant risk factor for malignant transformation of the oral epithelium and that bacterial interactions may contribute to OSCC development [14, 20]. Recent investigations suggest that a comprehensive understanding of the bacterial microbiome, its dynamics, and its alterations during health and disease could elucidate the microorganisms differentially associated with OSCC [14, 21]. Most research on the relationship between oral microbiota and cancer has been based on 16S rRNA gene sequencing. However, this methodology presents several limitations in terms of sensitivity, depth, and robustness, which remain major challenges in this field. In recent years, the development and application of high-throughput sequencing technologies (NGS) have enabled the analysis of complete metagenomes, allowing for more accurate bacterial identification at the species level, as well as a better understanding of their functional activity in various biological processes [34-36]. Therefore, it is essential to investigate the bacterial metagenome present in supragingival dental plaque, tumor tissue, and saliva from patients with and without OSCC.
T This study characterized the bacterial microbiota profiles associated with periodontopathogenic and cariogenic bacteria in patients with and without OSCC across two oral microenvironments. The diversity of both bacterial profiles varied between the two patient groups, with differences in species frequency and abundance. A total of 22 genera and 46 species were identified. The periodontopathogenic bacterial profile was characterized by the presence of 16 genera (e.g., Prevotella, Porphyromonas, and Fusobacterium) and 20 species, whereas the cariogenic bacterial profile was dominated by 6 genera (e.g., Streptococcus and Veillonella) and 26 species. Notably, the overall abundance of these bacterial profiles was low in both patient groups. Participants in the control group exhibited significantly better oral hygiene compared to the patient group (80% vs. 20%), whereas patients with OSCC demonstrated poor oral hygiene (60%) compared to the control group (20%).
Adequate oral hygiene is essential to prevent biofilm formation on tooth surfaces. Dental caries result from microbial dysbiosis within dental biofilms, leading to enamel and dentin demineralization due to bacterial acid production. Persistent biofilm accumulation can induce subgingival dysbiosis, triggering host immune responses and promoting periodontal disease [37]. Moreover, chronic inflammation has been strongly associated with malignant transformation of the oral epithelium. Oral bacteria can modulate the inflammatory microenvironment and interfere with host signaling pathways that regulate cell viability, proliferation, and differentiation, thereby contributing to the development of oral cancer [38-40].
In the present study, tumor localization was predominantly in the tongue (50%), followed by the cheeks (30%) and lips (20%). These results are consistent with those reported by Wolf et al. [22], who demonstrated a higher prevalence of tumors in the base of the tongue, floor of the mouth, cheeks, and soft palate among OSCC patients. Li et al. [41] assessed the microbial composition of plaque and saliva samples from OSCC patients and found tumors predominantly in the dorsal tongue and buccal mucosa. Similarly, Vesty et al. [42] reported a higher prevalence of tumors in the tongue (40%), cheeks (40%), and floor of the mouth (20%) among OSCC patients. Brian et al. [43] also found a high prevalence of tumors in the tongue (40%) and floor of the mouth (13.3%) in their study of the oral microbiome in cancer tissue samples.
The construction of the periodontopathogenic and cariogenic bacterial profiles was informed by a comprehensive literature review of studies employing DNA sequencing of subgingival plaque and saliva samples from patients with periodontal disease and caries. A total of 140 and 111 bacterial species were identified as potential contributors to periodontitis and caries, respectively.
It was also the intention of the research group to incorporate into the two profiles bacteria whose importance in periodontal disease and dental caries is not yet well established. In the periodontopathogenic bacterial profile, the following bacteria were incorporated: S. marcescens, P. aeruginosa, K. pneumoniae, H. parainfluenzae, and Aggregatibacter aphrophilus. These are Gram-negative, facultatively anaerobic bacteria possessing lipopolysaccharides, some with demanding growth characteristics that interact in the oral ecosystem with obligate anaerobes such as P. gingivalis, F. nucleatum, P. intermedia, and others [44]. Within the cariogenic profile, we also included, in addition to the classic cariogenic bacteria, different species of Streptococcus and Lactobacillus, whose association with dental caries is not yet fully defined and that, in some cases, are part of the normal oral microbial ecology of plaque and saliva [44].
Different studies have indicated the identification of periodontopathogenic, cariogenic, and other bacteria with specific characteristics in cancer patients based on gender. In the study by Chen et al. [45], the genera Megasphaera, Aggregatibacter, Atopobium, Lautropia, Actinobacillus, Bulleidia, Catonella, Filifactor, Corynebacterium, TG5, Acholeplasma, Moryella, Butyrivibrio, Dialister, Peptococcus, and Cardiobacterium were found in abundance in healthy tissue, oropharyngeal OSCC, and dysplastic tissue. Al-Hebshi et al. [46] reported the identification of multiple genera in three OSCC patients, with Haemophilus, Neisseria, Prevotella, Fusobacteria, Streptococcus, Porphyromonas, Leptotrichia, and Aggregatibacter being the most abundant. A separate study identified 120 genera in saliva samples, with Streptococcus and Prevotella being the most prevalent. Notably, Lactobacillus was significantly more abundant in head and neck squamous cell carcinoma compared to controls, while Citrobacter was most prevalent in oropharyngeal squamous cell carcinoma [24]. Wolf et al. [22] reported a higher prevalence of Streptococcus, Prevotella, and Rothia in both OSCC patients and healthy controls. Notably, Prevotella, Haemophilus, Neisseria, Streptococcus, and Veillonella were more abundant in healthy individuals, whereas Actinomyces, Schwartzia, Treponema, and Selenomonas were more prevalent in cancer patients. In a separate study, Dong et al. [23] identified Streptococcus, Neisseria, Prevotella, Actinobacillus, and Veillonella as the most abundant genera in samples from the oral cavity and esophagus.
Studies investigating bacterial species in OSCC at the species level are limited. Al-Hebshi et al. [47] identified Prevotella oris, Neisseria species, Fusobacterium nucleatum subsp. polymorphum, Aggregatibacter segnis, Streptococcus mitis, Fusobacterium periodonticum, Porphyromonas sp. oral taxon 279, and Alloprevotella tannerae as the predominant species in three OSCC patients. A subsequent study revealed F. nucleatum subsp. polymorphum as the most significantly expressed species in OSCC tumors, followed by Pseudomonas aeruginosa and Campylobacter sp. Oral taxon 44. In contrast, Streptococcus mitis, Rothia mucilaginosa, and Haemophilus parainfluenzae were significantly more abundant in control samples [22].
A recent study aimed to characterize the oral microbiome and cytokine levels in saliva from OSCC patients and healthy individuals through 16S rRNA sequencing of the V3/V4 regions using Illumina's MiSeq and cytokine analysis. Findings indicated a high abundance of Prevotella melaninogenica, Fusobacterium sp., Veillonella parvula, Porphyromonas endodontalis, Prevotella pallens, Dialister, Streptococcus anginosus, Prevotella nigrescens, Campylobacter ureolyticus, Prevotella nanceiensis, and Peptostreptococcus anaerobius, along with a significant increase in pro-inflammatory cytokines in the saliva of OSCC patients. Oncobacteria, including S. anginosus, V. parvula, P. endodontalis, and P. anaerobius, have been implicated in OSCC development through the upregulation of inflammatory cytokines such as IL-6, IL-8, TNF-α, IFN-γ, and GM-CSF [48]. However, Lactobacillus helveticus has been shown to possess immunomodulatory and anti-inflammatory properties, as it enhances immune function, reduces inflammation, and strengthens the intestinal mucosa [49]. Additionally, this eubiotic species has been reported to prevent infections by inhibiting pathogen adhesion and promoting bacterial aggregation and co-aggregation, thereby contributing to the competitive exclusion of pathogens. These mechanisms may significantly influence the composition and abundance of the microbiome [49, 50].
In this research, it is important to note the homogeneous distribution of the 26 bacterial species of the cariogenic profile in both study groups. In 12 of the 26 bacteria of the cariogenic profile, there was a high similarity in frequency (>80%) in both saliva and dental plaque samples in patients with and without OSCC. In the other 14 bacteria of the cariogenic profile (S. parasanguinis, S. constellatus, S. anginosus, R. mucilaginosa, L. salivarius, S. mutans, S. macedonicus, S. lutetiensis, L. helveticus, L. reuteri, S. parauberis, S. infantarius, and S. pasteurianus), there were some differences in frequency between the two study groups. Of this last group, the bacterial species that were found with low frequency in the saliva of patients with OSCC and with high frequency in the saliva of patients without OSCC are noteworthy: L. helveticus (10% versus 60%), L. reuteri (20% versus 80%), S. pasteurianus (30% versus 80%), S. parauberis (30% versus 80%), S. infantarius (30% versus 80%), S. lutetiensis (40% versus 80%), S. macedonicus (50% versus 90%), and Enterobacter asburiae (0% in patients with OSCC and 50% in patients without OSCC). Currently, worldwide research is underway to identify bacterial biomarkers, as they could be useful for monitoring the course of some types of cancer and designing strategies for its prevention and control.
In this sense, it has been proposed that differentially abundant microorganisms could be used as biomarkers of pathological or healthy states [51]. Very likely, these eight bacterial species found with greater frequency and statistically significant differences in cancer-free patients could serve as cancer biomarkers if the internal parameters and conditions within the oral cavity in which they develop and are part of the normal microbial flora can be clearly established, and if, for some reason, they are found less frequently in cancer patients [21, 51]. Possibly, at some point, these bacteria will be recognized as eubiotic and become candidates for detection by PCR amplification systems. They could also serve as markers of oral health when cultured from saliva or plaque of cancer-free patients.
The distinct microbial community structures observed in the PCoA analysis suggest a potential association between OSCC status and microbiome composition, although the small sample size limits the generalizability of these findings.
Regarding the periodontopathogenic bacterial profile, the main periodontopathogenic bacteria: P. denticola, T. forsythia, P. melaninogenica, P. intermedia, P. gingivalis, H. parainfluenzae, F. nucleatum, C. ochracea, A. aphrophilus, and A. actinomycetemcomitans were found at high frequency (>80%) and were homogeneously distributed in both types of samples in both study groups. On the other hand, due to their low frequency in saliva and absence in dental plaque, the bacteria P. stuartii, G. vaginalis, and C. canimorsus stand out in patients without OSCC. As a potentially eubiotic bacterium, E. asburiae stands out in saliva and dental plaque samples from patients without OSCC (50%), in contrast to its absence in saliva from patients with OSCC.
Among the studies describing bacterial species in oral cancer, Al-Hebshi and collaborators 2015 [47] identified 228 bacterial species in three patients with OSCC. The most abundant were Prevotella oris, Neisseria flava, N. flavescens/subflava, Aggregatibacter segnis, Streptococcus mitis, Porphyromonas sp. oral taxon 279, Fusobacterium periodonticum, Fusobacterium nucleatum ss polymorphum Neisseria elongata, and Alloprevotella tannerae. The species Streptococcus infantarius, Streptococcus lutetiensis, Streptococcus macedonicus, Streptococcus pasteurianus and Streptococcus parauberis, proposed as biomarkers of eubiosis, have not been described in OSCC; however, they have been reported in other types of cancer such as colorectal cancer (Streptococcus macedonicus) [52], colon carcinoma (Streptococcus infantarius) [53, 54], colon cancer (Streptococcus pasteurianus) [55], hepatocellular carcinoma (Streptococcus pasteurianus) [56], and malignant and premalignant colonic lesions (Streptococcus pasteurianus) [57].
In contrast, species such as Streptococcus mitis, Streptococcus taxon 070 and 058, Streptococcus sanguinis, Streptococcus dentisani, Streptococcus australis, and Streptococcus cristatus have been found more frequently and abundantly in healthy individuals compared to OSCC patients [58]. Some studies have proposed Streptococcus mitis, Streptococcus anginosus, Streptococcus sobrinus, and Streptococcus salivarius as potential diagnostic and prognostic biomarkers for cancer [1, 39].
Regarding the biological significance of these bacterial species with potential eubiotic properties, Lactobacillus helveticus has been described as having immunomodulatory and anti-inflammatory properties, as it enhances intestinal immune function, reduces inflammation, and strengthens the intestinal mucosa. Additionally, it has been reported that the eubiotic species Lactobacillus helveticus has the ability to prevent infections by inhibiting pathogen adhesion and promoting bacterial aggregation and co-aggregation, thereby contributing to the competitive exclusion of pathogens, which could effectively impact the composition and abundance of any microbiome [49]. Lactobacillus reuteri, in turn, has been associated with the maintenance of oral health, as it reduces gingival inflammation and dental plaque formation, thereby lowering the risk of caries [50]. Although Enterobacter asburiae belongs to a group that includes opportunistic pathogens, some of its strains have demonstrated the ability to produce antimicrobial compounds and compete with pathogenic bacteria in the gut. However, its use as a probiotic remains controversial and requires further investigation [59]. On the other hand, the protective role of non-pathogenic or commensal Streptococcus species is not well documented. However, some studies suggest that S. macedonicus may produce bacteriocins with antimicrobial activity and has been isolated from fermented dairy products, indicating a possible probiotic role. S. infantarius and S. lutetiensis have been identified in healthy human microbiotas, although their specific protective functions are still under investigation. S. parauberis and S. pasteurianus have been studied mainly in veterinary contexts, although some studies have detected them in human microbiotas without clear pathological associations [60].
The human oral microbiome is a dynamic microbial community whose composition changes over time, influenced by physiological, environmental, and behavioral factors. From birth to old age, there is a progressive evolution in the diversity and abundance of bacterial species, modulated by events such as tooth eruption, hormonal changes, diet, oral hygiene, and medication use. Factors such as high sugar intake, poor oral hygiene, prolonged antibiotic use, and systemic conditions like diabetes can induce dysbiosis, disrupting microbial balance and promoting the onset of oral and systemic diseases. This knowledge highlights the importance of monitoring and modulating the oral microbiome throughout the life cycle as an integral part of promoting both oral and general health [61]. Analyzing how the composition of the oral microbiome varies in the early stages of the transition from health to disease may facilitate the identification of relevant prognostic markers and the development of new therapeutic strategies aimed at preventing or reversing disease progression before irreversible damage occurs [62-64].
Among the limitations of the present study, confounding variables such as host immunity-related factors, sex, oral hygiene, dental and periodontal status, nutritional habits, smoking, and alcohol consumption were considered. However, stratifying patients with OSCC according to their periodontal or caries status was not possible. Additionally, individual variations related to place of origin, diet, oral hygiene, smoking, and alcohol consumption could have influenced the results.
CONCLUSION
In the two study groups, the cariogenic and periodontopathogenic bacterial profiles were established, with a wide and variable distribution of bacteria in the plaque and saliva samples. The bacterial species L. helveticus, L. reuteri, S. pasteurianus, S. parauberis, S. infantarius, S. lutetiensis, S. macedonicus, and E. asburiae could have potential future eubiotic utility.
It is recommended that future studies include functional analyses to validate the roles of the proposed bacteria in eubiotic states.
AUTHORS’ CONTRIBUTIONS
The authors confirm their contributions to the paper as follows: A.E., F.O.G.J., D.A.G.R.: Conceptualization; A.E, F.T.T.: Data Curation; A.E.: Formal Analysis; A.E, D.A.G.R.: Investigation; A.E., D.A.G.R., F.O.G.J., A.E.: Methodology; D.A.G.R, F.O.G.J, A.E.: Supervision; A.E., F.T.T., D.A.G.R., F.O.G.J.: Validation; A.E., F.O.G.J., D.A.G.R.: Writing – Original Draft; All authors: Writing – Review & Editing.
LIST OF ABBREVIATIONS
| OSCC | = Oral Squamous Cell Carcinoma |
| PCoA | = Principal Coordinate Analysis |
| FASTQ | = format that stores DNA sequences along with information about the quality of each base. |
| HOMD | = Human Oral Microbiome Database |
| FDR | = False Discovery Rate |
| PERMANOVA | = Permutational Multivariate Analysis of Variance. |
| GM-CSF | = Granulocyte-Macrophage Colony-Stimulating Factor. |
| TNF-α | = Tumor Necrosis Factor alpha. |
| IFN-γ | = Interferon gamma. |
| PCR | = Polymerase Chain Reaction. |
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
The study was approved by the ethics committee of the Pontificia Universidad Javeriana, Bogotá D.C., Colombia with approval number OD-0149.
HUMAN AND ANIMAL RIGHTS
All procedures performed in studies involving human participants were in accordance with the ethical standards of institutional and/or research committee and with the 1975 Declaration of Helsinki, as revised in 2013.
AVAILABILITY OF DATA AND MATERIAL
Data supporting the findings of this article can be made available upon reasonable request.
FUNDING
This study was supported by the Universidad Cooperativa de Colombia, Bogotá, Funder ID: NIT 860.029.924-7 (Awards/Grant number: INV3148).
ACKNOWLEDGEMENTS
The authors express their gratitude to the Pontificia Universidad Javeriana, Bogotá, and the Universidad Cooperativa de Colombia, Columbia for funding the present study.

