All published articles of this journal are available on ScienceDirect.

Dissemination of Plasmid-Mediated Aminoglycoside-Modifying Enzymes Among MDR Acinetobacter baumannii Isolates from a Tertiary Care Egyptian Hospital
Authors Info & Affiliations
Abstract
Background:
Acinetobacter baumannii is one of the most challenging multidrug-resistant (MDR) nosocomial pathogens worldwide. Aminoglycosides are used for the treatment of A. baumannii infections, however, resistance to aminoglycosides is currently emerging, limiting therapeutic choices.
Objective:
In this study, the prevalence of aminoglycoside resistance and plasmid-mediated mechanisms of aminoglycoside resistance were investigated in A. baumannii clinical isolates collected from ICU patients at a tertiary care hospital in Egypt.
Methods:
The automated Vitek 2 system was used to identify A. baumannii species and determination of the antimicrobial susceptibility pattern. The identification of A. baumannii was confirmed by the detection of the blaOXA-51-like gene intrinsic to this species. Minimum Inhibitory Concentration (MIC) of gentamicin was determined using E-test following the CLSI breakpoints. Isolates were screened for the prevalence and diversity of the plasmid-carried aminoglycoside-modifying enzymes encoding genes aacC1, aadA1, aadB and aphA6. For genetic diversity analysis, the ERIC-PCR method was performed.
Results:
All A. baumannii isolates were MDR with high resistance rates to tested antimicrobials. The resistance rate to gentamicin was 92.9% with elevated MICs (≥ 32 μg/mL). The gentamicin-resistant isolates harboured one or more of the studied genes with the prevalence of aphA6 (81%). ERIC-based genotyping revealed that there was no evidence of A. baumannii clonal dissemination among isolates.
Conclusion:
The study concluded that MDR A. baumannii isolates were highly resistant to gentamicin. The plasmid-carried aminoglycoside-modifying enzymes encoding genes were disseminated among isolates with the AphA6 gene, which was the most prevalent one. The acquisition of more than one aminoglycoside resistance gene was associated with an elevated MIC of gentamicin. Thus, regular surveillance studies of the emerging resistance to antimicrobials and strict measures to control the dissemination of resistance determinants genes are warranted.
1. INTRODUCTION
A. baumannii is a gram-negative, nonfermentative, oxidase-negative coccobacillus that has emerged to an infectious agent of importance worldwide [1]. A. baumannii has been ranked second after Pseudomonas aeruginosa among Gram-negative pathogens causing infections in the hospitalized immunocompromised patients. This organism is responsible for a significant proportion of nosocomial infections, including urinary tract infections, surgical-site infections, meningitis, endocarditis, septicemia, and ventilator-associated pneumonia among the Intensive Care Unit (ICU) patients [2-8]. Moreover, A. baumannii has the ability to accumulate mechanisms of resistance to various classes of antimicrobials, leading to the emergence of Multidrug-Resistant (MDR) and Extensive Drug-Resistant (XDR) strains that are difficult to treat [1, 7, 9, 10]. Centers for Disease Control and Prevention (CDC) report highlighted MDR Acinetobacter as a serious threat that causes about 7,000 infections and about 500 deaths in the United States each year [11].
Antimicrobial agents of choice in treating A. baumannii infections include sulbactam, carbapenems, polymixins, tigecycline, tetracyclines and aminoglycosides. Combination therapy is usually required for effective treatment, particularly against MDR- and XDR-caused infections [12, 13]. Aminoglycosides have long been used as an alternative or synergistic agent with cell-wall inhibitors for the treatment of infections caused by MDR A. baumannii strains. However, in recent years, resistance rates have emerged and resistance mechanisms have increased against aminoglycosides, which may also impair synergism with other antimicrobials making it challenging to treat such infections [8, 14, 15].
Aminoglycoside resistance in Acinetobacter is attributed mainly to the production of Aminoglycoside-Modifying Enzymes (AMEs), including acetyltransferases (AACs), nucleotidyltransferases (ANTs) and/or phosphotransferases (APHs). Several AMEs have been detected in Acinetobacter spp. causing clinical resistance; particularly the presence of AACs and APHs enzymes, which can cause a high level of resistance. Genes encoding AMEs in A. baumannii can be located on mobile genetic elements, including plasmids, transposons, or class I integrons [6, 15-19]. The current study aimed to evaluate the prevalence of aminoglycoside resistance; clonal dissemination; and the occurrence rate of the common plasmid-carried aminoglycosides resistance encoding genes, aacC1 (encodes gentamicin acetyltransferase I or aminoglycoside N(3')-acetyltransferase I), aadA1 (encodes aminoglycoside nucleotidyltransferase), aadB (encodes aminoglycoside (2') adenyltransferase) and aphA6 (aminoglycoside 3'-phosphotransferase), among A. baumannii isolates from patients admitted to the ICU at a tertiary hospital in Cairo, Egypt.
2. MATERIALS AND METHODS
2.1. Clinical Isolates and Identification of A. baumannii
A total of 85 non-duplicate A. baumannii clinical isolates were included in this study, collected from the tertiary hospital, International Medical Centre (IMC), Cairo, Egypt, upon the approval of the included hospital of study. These 85 A. baumannii isolates were recovered from various clinical samples, collected from patients admitted to ICUs between 2015 and 2018, including Central Venous Pressure (CVP) catheter tips (14/85, 16.47%), endotracheal tube aspirate (29/85, 34.12%), wound swabs (22/85, 25.88%), urine (5/85, 5.88%), blood (12/85, 14.18%) and sputum (3/85, 3.53%). These samples were routinely collected and processed by the dedicated team during the medical care of ICU patients. A. baumannii isolates were identified by the Vitek 2 automated system (bioMe´rieux, France) at IMC. The identification of A. baumannii was further confirmed by the detection of the intrinsic blaOXA-51-like gene by polymerase chain reaction (PCR) [20].
2.2. Antimicrobial Susceptibility Testing and Determination of Gentamicin MIC
The susceptibility of A. baumannii isolates to 16 diverse antimicrobials was determined by Vitek 2. The gentamicin MIC was further determined on gentamicin-resistant isolates by E-test (bioMérieux) on Mueller-Hinton agar following the manufacturer's instructions. Concentrations of gentamicin in strip (GM) ranged from 0.064 - 1024 μg/mL. The results were interpreted according to CLSI (M100-28th edition) breakpoints [21]: MIC ≤ 4 μg/mL for susceptible and MIC = 8 for intermediate and MIC ≥ 16 μg/mL for resistant. Escherichia coli ATCC 25922 was used as a quality control strain. The A. baumannii isolate that showed resistance to at least three or more different antimicrobial classes was considered as MDR [13, 19]. The frequencies and percentages were used to present antimicrobial resistance profiles and the distribution of resistance determinants encoding genes.
2.3. PCR Amplification of the Resistance Genes blaOXA-51-like, aacC1, aadA1, aadB and aphA6
The blaOXA-51-like carbapenemase gene (GenBank accession no. NZ_CP020596.1) and AMEs plasmid-carried genes aacC1 (GenBank accession no. KR610434.1), aadA1 (GenBank accession no. NC_011586.2), aadB (GenBank accession no. NC_010410.1) and aphA6 (GenBank accession no. NZ_CP023030.1) in A. baumannii isolates were amplified by PCR. PCR oligonucleotide primers used in this study, synthesized by Invitrogen, UK, are listed in Table 1. Total DNA was extracted from all tested isolates using the boiling method by heating bacterial cells suspension in sterile distilled water at 95°C for 10 min, followed by the removal of cellular debris by centrifugation at 14,000 rpm for 1 min. The supernatant was collected and used as template DNA for PCR amplification. PCR reaction mixtures were prepared in total volumes of 20 μl. Each reaction contained 1 μl of template DNA, 1 μl (equivalent to 10 pmol concentration) of each primer and 10 μl of GoTaq® Green Master 2× Ready Mix (Promega, USA), then the volume was completed to 20 μl by adding 7 μl of nuclease-free water. The PCR amplification programs were as follows: initial denaturation for 5 min at 95°C, then 30 cycles of denaturing at 95°C for 30 seconds, annealing for 30 seconds at 52°C for blaOXA-51-like, 58°C for the aacC1 gene, 55°C for the aadA1 gene, aadB gene and aphA6 gene and extension at 72°C for 45 seconds, followed by a final extension at 72°C for 7 min.
| Target gene | Primer sequence (5' – 3') | Expected amplicon size | Source |
|---|---|---|---|
| blaOXA-51 | F: TAATGCTTTGATCGGCCTTG R: TGGATTGCACTTCATCTTGG |
353 bp | [20] |
| aacC1 | F: GCAGTCGCCCTAAAACAAAG R: CCCGTATGCCCAACTTTGTA |
456 bp | [17] |
| aadA1 | F: GAAGCCACACAGTGATATTGATTT R: GTTCCATAGCGTTAAGGTTTCATT |
397 bp | [22] |
| aadB | F: GCATATCGCGACCTGAAAGC R: GACACAACGCAGGTCACATTGAT |
524 bp | [17] |
| aphA6 | F: ATGGAATTGCCCAATATTATTC R: TCAATTCAATTCATCAAGTTTTA |
780 bp | [23] |
2.4. Enterobacterial Repetitive Intergenic Consensus Polymerase Chain Reaction (ERIC-PCR) Genotyping and Analysis of ERIC Patterns
ERIC-PCR-based genotyping was carried out to determine the genetic diversity among A. baumannii isolates [22, 23-27]. Genomic DNA was extracted from A. baumannii isolates using commercially available GeneJET Genomic DNA purification Kit (Thermo Scientific, USA). The PCR reactions were prepared in total volumes of 25 μl, contained ~10 ng of template DNA, 10 pmol of ERIC-2 primer (5'-AAGTAAGTGACTGGGGTGAGCG -3'), 12.5 μl of Dream Taq Green PCR Master Mix (2X) (Promega, USA) and the volume was completed to 25 μl by adding nuclease-free water. The amplifications were performed in Veriti 96 well Thermal Cycler (Applied Biosystems, USA) programmed for an initial denaturation at 94°C for 5 mins and 35 cycles of denaturation at 94°C for 1 min, primer annealing at 45°C for 1 min and extension at 72°C for 2 mins, followed by a final extension at 72°C for 7 mins. The obtained ERIC patterns were clustered by dendrogram generated with the Dice similarity coefficient and the UPGMA clustering method using DendroUPGMA software (http://insilico.ehu.es/dice_upgma/).
2.5. TAE (Tris-acetate-EDTA)-Agarose Gel Electrophoresis
PCR amplified products and ERIC-PCR patterns were subjected to TAE agarose gel (1% gel stained with ethidium bromide) (Bioline, UK) electrophoresis. GeneRuler 1 kb DNA molecular weight marker (Thermo Scientific, USA) was used for sizing the PCR products. DNA fragments were visualized by placing on a UV transilluminator (UVP, LLC, UK) and photographed directly.
3. RESULTS
3.1. Identification and Antimicrobial Resistance Patterns of A. baumannii Isolates
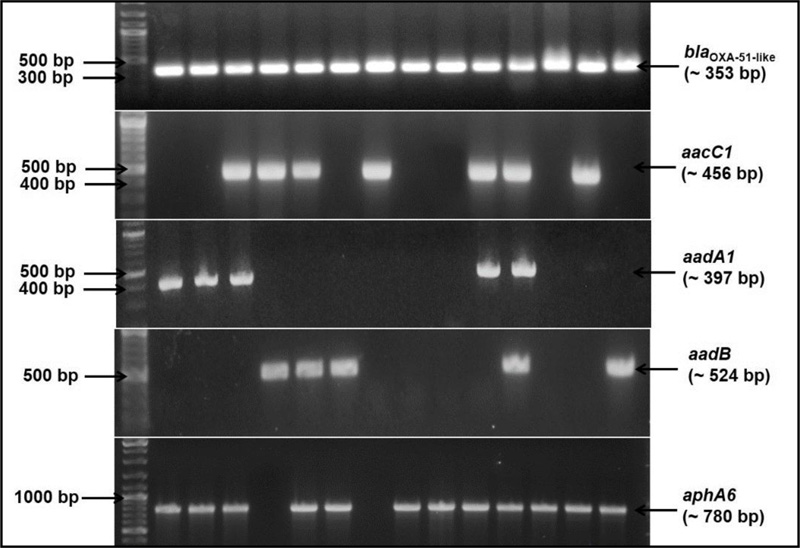
Vitek 2 identified A. baumannii species with a level of 99% probability. A. baumannii intrinsic blaOXA-51-like gene was amplified from all 85 isolates (Fig. 1). Antimicrobial susceptibility testing indicated that all 85 A. baumannii isolates (100%) were MDR. All the isolates were resistant to ampicillin, ampicillin-sulbactam, piperacillin-tazobactam, cefuroxime, cefuroxime axetil, cefpodoxime, cefotaxime, ceftazidime, imipenem, meropenem and ciprofloxacin antimicrobials. Resistance rates were also high against other antimicrobials, including moxifloxacin (92.9%), trimethoprim-sulfamethoxazole (90.6%), tetracycline (73%) and gentamicin (88.2%). Tigecycline showed the highest activity against A. baumannii isolates; its resistance rate was 0% (Table 2).
| Antimicrobial agent |
Sensitive (S) No. (%)* |
Intermediate (I) No. (%)* |
Resistant (R) No. (%)* |
|---|---|---|---|
| Ampicillin | 0 (0) | 0 (0) | 85 (100) |
| Ampicillin/Sulbactam | 0 (0) | 0 (0) | 85 (100) |
| Piperacillin/Tazobactam | 0 (0) | 0 (0) | 85 (100) |
| Cefuroxime | 0 (0) | 0 (0) | 85 (100) |
| Cefuroxime Axetil (Ceftin) | 0 (0) | 0 (0) | 85 (100) |
| Cefpodoxime | 0 (0) | 0 (0) | 85 (100) |
| Cefotaxime | 0 (0) | 0 (0) | 85 (100) |
| Ceftazidime | 0 (0) | 0 (0) | 85 (100) |
| Imipenem | 0 (0) | 0 (0) | 85 (100) |
| Meropenem | 0 (0) | 0 (0) | 85 (100) |
| Ciprofloxacin | 0 (0) | 0 (0) | 85 (100) |
| Moxifloxacin | 0 (0) | 6 (7.1) | 79 (92.9) |
| Gentamicin | 6 (7.1) | 4 (4.7) | 75 (88.2) |
| Tetracycline | 7 (8.2) | 16 (18.8) | 62 (73) |
| Tigecycline | 56 (65.9) | 29 (34.1) | 0 (0) |
| Trimethoprim/Sulfamethoxazole | 10 (9.4) | 0 (0) | 77 (90.6) |
| AMEs encoding genes | Gentamicin-resistant A. baumannii phenotypes (79 isolates) |
|---|---|
| Genotypic profile | No. (%)* |
| aphA6 | 64 (81%) |
| aacC1 | 42 (53.2%) |
| aadA1 | 43 (51.9%) |
| aadB | 28 (35.4%) |
| AME single-producer gentamicin-resistant A. baumannii (19 isolates) | |
| Genotypic profile | No. (%)** |
| aacC1 alone | 2 (10.5) |
| aadA1 alone | 1 (5.3) |
| aadB alone | 1 (5.3) |
| aphA6 alone | 15 (78.9) |
| AMEs co-producer gentamicin-resistant A. baumannii (60 isolates) | |
| Genotypic profile | No. (%)*** |
| aacC1 + aadA1 + aadB + aphA6 | 5 (8.3) |
| aacC1 + aadA1 + aphA6 | 24 (40) |
| aadA1 + aadB + aphA6 | 2 (3.3) |
| aacC1 + aadA1 | 8 (13.4) |
| aadA1 + aphA6 | 3 (5) |
| aadB + aphA6 | 10 (16.7) |
| aadB + aphA6 | 5 (8.3) |
| aacC1 + aadB | 3 (5) |
3.2. The MIC of Gentamicin to A. baumannii Isolates Using E-test
The MIC of gentamicin against A. baumannii isolates, determined by E-test, revealed that 7.1% (6/85) of isolates were susceptible to gentamicin with MICs ≤ 4 μg/mL. The majority of isolates, 92.9% (79/85), were resistant to gentamicin with MICs ranged from 16 to ≥ 512 μg/mL (Table 4).
3.3. Distribution of Plasmid-Carried Aminoglycoside Resistance Encoding Genes
The genotypic profile of the 79 gentamicin-resistant A. baumannii isolates, phenotypically confirmed as AME-producer by E-test, was investigated. In general, PCR-based molecular study revealed the presence of one or more of AMEs encoding genes in all investigated isolates. The gene aphA6 was the most predominant gene carried by these gentamicin-resistant A. baumannii phenotypes, then aacC1, aadA1 and aadB with frequencies 81% (64/79), 53.2% (42/79), 51.9% (43/79) and 35.4% (28/79), respectively (Table 3 and Fig. 1). None of these genes was found in the six gentamicin-sensitive isolates. Almost 24% (19/79) of these isolates harboured only one of the resistance genes. Most of these single-producers, 78.9% (15/19), were aphA6 positive. However, 75.9% (60/79) of isolates were co-producers of AMEs encoding genes indicated by either two, three or all the four investigated genes were co-existed together in the same isolate. Of the 60 co-producer isolates, 8.3% were positive for the four resistance genes aacC1, aadA1, aadB and aphA6, 43.3% of isolates were positive for three genes and 48.4% of isolates were positive for two of the tested genes in different combinations. The most prevalent genotypic profile was the aacC1, aadA1 and aphA6 together; it was found in 40% of the AMEs co-producers isolates (Table 3).
3.4. Correlation of MICs of Gentamicin with the Presence of AMEs Encoding Genes
Correlation between MIC and the presence of AMEs encoding genes showed that the highest frequency of resistant isolates, 89% (58/65), that have MIC ≥ 32 μg/mL harboured more than one of the AMEs encoding genes. However, 86% (12/14) that have MIC 8 to 16 μg/mL were positive for only one of the AMEs encoding genes. As expected, none of the 6 (0%) susceptible isolates harboured any of the AMEs genes (Table 4).
3.5. Gentamicin-resistant A. baumannii Isolates were Polyclonal
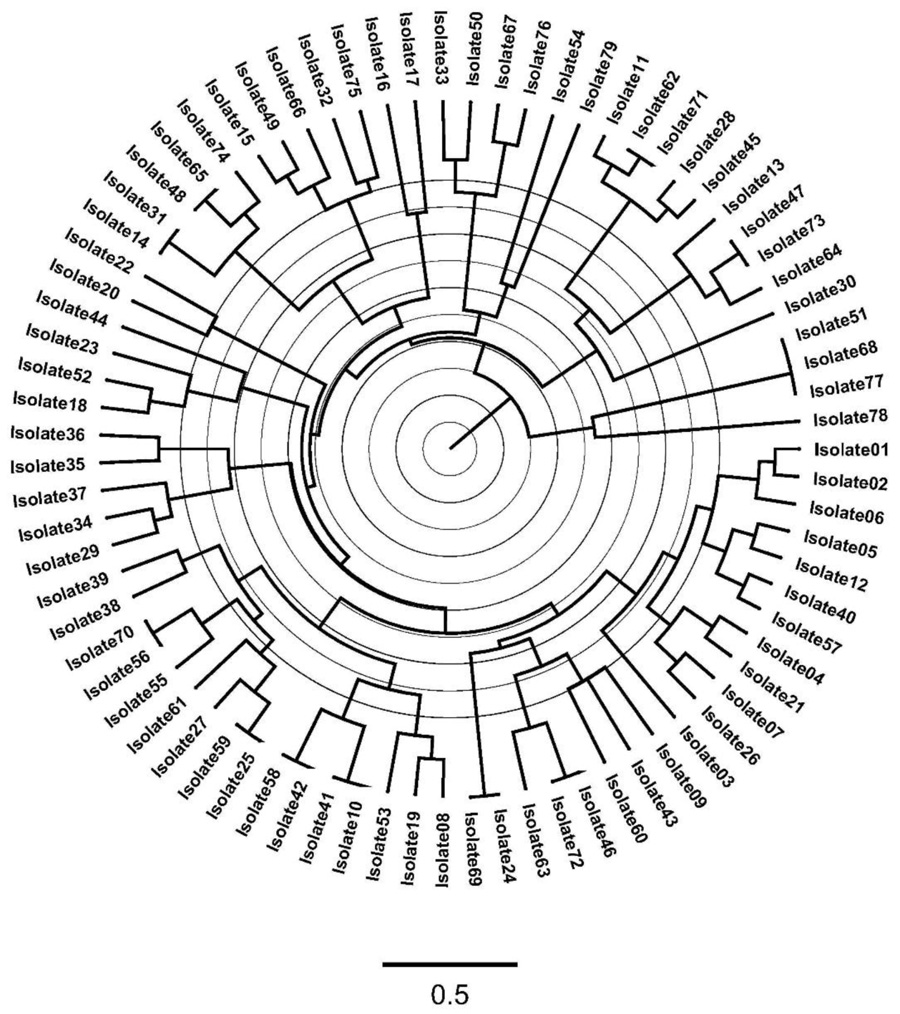
ERIC-PCR-based genotyping of the 79 A. baumannii isolates revealed a significant molecular heterogeneity among isolates from the hospital of study with a small percentage, 15% of isolates showed closer genetic relatedness, which is indicated by 67 different ERIC-based patterns or fingerprints (Fig. 2).
| Resistance pattern | Distribution of MIC of gentamicin (µg/mL)* | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| No. (%) | |||||||||
| R | S | ≤ 4 | 8 | 16 | 32 | 64 | 128 | 256 | ≥ 512 |
| 79 (92.9%) | 6 (7.1%) | 6 | 0 | 10 | 18 | 22 | 14 | 6 | 9 |
| Genotypic profile | |||||||||
| aacC1 alone (2)** | - | - | - | - | 2 | - | - | - | |
| aadA1 alone (1) | - | - | - | 1 | - | - | - | - | |
| aadB alone (1) | - | - | 1 | - | - | - | - | - | |
| aphA6 alone (15) | - | - | 7 | 4 | - | - | 4 | - | |
| aacC1 + aadA1 + aadB + aphA6 (5) | - | - | - | - | 2 | 1 | 2 | - | |
| aacC1 + aadA1 + aphA6 (24) | - | - | - | 1 | 12 | 2 | - | 9 | |
| aadA1 + aadB + aphA6 (2) | - | - | - | 2 | - | - | - | - | |
| aacC1 + aadA1 (8) | - | - | 2 | - | - | 6 | - | - | |
| aadA1 + aphA6 (3) | - | - | - | 3 | - | - | - | - | |
| aadB + aphA6 (10) | - | - | - | 7 | - | 3 | - | - | |
| aadB + aphA6 (5) | - | - | - | - | 4 | 1 | - | - | |
| aacC1 + aadB (3) | - | - | - | - | 2 | 1 | - | - | |
4. DISCUSSION
A. baumannii infection is one of the most important healthcare problems encountered in hospitals, clinics and public health centers [18]. The inherent and acquired resistance against multiple antimicrobial classes makes it difficult to treat A. baumannii-caused infections, particularly those caused by MDR and XDR strains [6, 25]. Aminoglycosides have been an important group of antibiotics commonly used in the treatment of serious A. baumannii-caused infections either alone or in synergistic combination with other antimicrobials. However, recent reports indicated the emergence of resistance to aminoglycosides in Acinetobacter spp. worldwide. Aminoglycoside resistance in Acinetobacter is primarily mediated by the productions of plasmid-mediated AMEs [6, 15, 18, 26]. Therefore, this study focused on exploiting the resistance pattern to gentamicin, one of the frequently used aminoglycosides, in clinical isolates of A. baumannii. The prevalence of plasmid-carried AMEs encoding genes and the clonal variation of the clinical isolates were also investigated. All 85 isolates included in this study were confirmed as A. baumannii by the PCR-based detection of the intrinsic blaOXA-51, which can distinguish A. baumannii accurately from other members of the Acinetobacter genus [17]. Vitek 2 identification results were totally agreed to the more reliable blaOXA-51-based genetic identification, thus, providing a rapid and accurate method for the identification of A. baumannii, particularly in the overcrowded healthcare settings [20, 28].
Following the definitions of multi-drug resistance and extensively drug-resistance by Magiorakos et al., antimicrobial susceptibility patterns revealed that all A. baumannii isolates were considered as MDR, with the possibility of being extensively drug-resistant (XDR) [29]. In this study, high resistance rates were recorded against all tested antimicrobials, however, only tigecycline showed good activity against A. baumannii isolates. This high resistance profile could be attributed to the possibility of being of nosocomial origin. In addition, the antimicrobial resistance records in this study are regrettably indicating that the tested antimicrobials are not suitable as empirical therapy for A. baumannii-caused infection in a developing country like Egypt, which makes the treatment challenging. This could be due to the overuse and/or misuse and the hospitals' routine use of these antimicrobials [30]. However, the effectiveness of tigecycline in the current study makes it as a potential alternative therapy for A. baumannii infections.
The rates of resistance to diverse antimicrobials among MDR A. baumannii isolates in this study were significantly higher than the rates recently recorded in Egypt, particularly against aminoglycosides [31, 32]. Noticeably, the incidence rate of resistance to aminoglycosides and /or gentamicin in the current study was nearly double the rates reported in these studies, indicating the emergence of resistance to aminoglycosides in Egypt. This could be explained by the extensive use of this class of antimicrobials, particularly gentamicin, during the latter years. An almost similar rate of gentamicin resistance (86%) to our study was reported by Aliakhbarzade et al. in Iran [6]. Significantly, several previous reports, from different geographical regions worldwide, have agreed to our findings as they have documented high rates of antimicrobial resistance in A. baumannii and showed that this pathogen is frequently resistant to multiple antimicrobial agents, with the emergence of strains that are resistant to almost all clinically relevant antimicrobials. Although, in these studies, different antimicrobial susceptibility patterns of A. baumannii have been observed between different countries, which likely can be attributed to environmental factors and different patterns and guidelines of antimicrobial usage [13, 16, 33, 34]. Owing to the limitations of the Vitek 2 system in antimicrobial susceptibility testing of the clinically important bacteria, it would be better in clinical microbiology and research to evaluate our own MICs [28]. Accordingly, in the current study, the susceptibility of isolates to gentamicin and gentamicin MIC were determined using the agar E-test method. E-test results indicated the resistance rate to gentamicin is 92.9%, which is more reliable, however, no intermediate activity was shown by the Vitek 2 system.
AMEs catalyse the modifications at -OH or -NH2 groups of the 2-deoxystreptamine nucleus or the sugar moieties and can be acetyltransferases (AACs), nucleotidyltransferases (ANTs), or phosphotransferases (APHs). In this study, 85 A. baumannii isolates were screened for plasmid-carried AMEs resistance genes by PCR. One of the AACs encoding genes (aacC1), two of the ANTs encoding genes (aadA1 and aadB1), and one of APHs encoding genes (aphA6) were selected for this screening given that these genes were found to be prevalent amongst aminoglycoside-resistant Acinetobacter spp. and other Gram-negative pathogens such as E. coli and Klebsiella spp. as well [4, 34, 35]. These genes were not found in the 6 gentamicin-susceptible isolates. However, one or more of the tested AMEs encoding genes were found in the 79 gentamicin-resistant genes. AphA6 was the most predominant detected gene in gentamicin resistant A. baumannii isolates (81%), followed by both aacC1 (53.2%) and aadA1 (51.9%). aadB gene was the lowest prevalent as it was found in only 35.4% of the resistant isolates. Previous reports recorded the prevalence of accC1 was 63.3% and 65.11%, respectively, which was higher than the rate reported in the current study [6, 13]. However, the incidence rates of AphA6, aadA1 and aadB were significantly higher in the current study. Lee et al. (2011) showed very comparable incidences rates of AMEs genes to our study; aacC1 (56%), aadB (48%), and aphA6 (71%) [36].
In the current study, 75.9% (60/79) of gentamicin-resistant isolates were positive for two or more of tested AMEs plasmid-carried genes. Of the 60 co-producer isolates, 5 strains (8.3%) were quadruple-positive for all tested genes aacC1, aadA1, aadB and aphA6. The most prevalent genotypic profile was the triple-positive aacC1, aadA1 and aphA6 profile; it was positive in 40% of the co-producer isolates. 48.4% were double-positive for two of the tested genes in different combinations; aadA1 + aphA6, aadB + aphA6, aadB + aphA6, or aacC1 + aadB. The aacC1 and aphA6 genes in the co-producer strains are the most common AME genes in A. baumannii isolates in agreement with previous reports [6, 13, 36]. Interestingly, we found that most of the isolates that have higher MICs (MIC ≥ 32 μg/mL) harboured more than one of the AMEs encoding genes, however, most of the isolates with MIC 16 μg/mL were positive for only one of the AMEs genes. This indicates that the presence of more than one gene of the aminoglycosides resistance genes not only increases the incidence rate of resistance to gentamicin but also elevates the level of MIC of this antimicrobial.
Genotypic diversity among the 79 gentamicin-resistant A. baumannii was analyzed using ERIC-PCR-based genotyping, which may help in the epidemiological investigation and knowing more about the bacteria acquisition of resistance determinants and transmission [37]. The ERIC data showed that there was significant molecular heterogeneity among these isolates collected from ICU patients at tertiary IMC hospital. Isolates were clustered into distinct genotypes and no specific clone dominated with a very low percentage of genetic relatedness. These findings indicated that bacteria were not transmitted between patients in ICU, in addition, there was no common source of infection as an exogenous source, which is very important from nosocomial and infection control standpoints. Moreover, this high clonal diversity of isolates revealed the dissemination of aminoglycosides resistance encoding genes in A. baumannii bacteria.
CONCLUSION
The study concluded that A. baumannii-caused infections remain difficult to treat and is considered a real health problem in Egypt as well as other regions of the world. A. baumannii bacteria have become highly resistant to diverse antimicrobial agents; including aminoglycosides. In the current study, we found that 93% of MDR A. baumannii isolates were resistant to aminoglycosides (i.e., gentamicin) with the majority of MICs ≥ 32 μg/mL; which is alarming as the therapeutic options will be limited. These high resistance rates may be consequent to over and misuse antimicrobials in developing countries like Egypt, thus, a public health policy on proper prescribing and the rational use of antimicrobials as well as developing new antimicrobials is needed. AphA6 was the most prevalent AMEs plasmid-mediated gene. The carriage of more than one AMEs encoding gene was associated with high-level gentamicin resistant phenotype indicated by elevated MIC. Although there was no evidence of clonal distribution between the tested A. baumannii isolates, strict infection control guidelines should be adopted, particularly in hospitals with a high incidence of A. baumannii infections to avoid the dissemination of resistance determinants among bacteria, such as transferable plasmid-carried AMEs resistance genes. In future studies, we will assess the effect of combination therapy of aminoglycosides with different antimicrobials to evaluate possible synergistic effects to overcome the gentamicin resistance in A. baumannii isolates, particularly those found to be AMEs co-producers. In addition, regular surveillance studies of the emerging resistance to antimicrobials are necessary.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
Not applicable.
HUMAN AND ANIMAL RIGHTS
No animals/humans were used for studies that are the basis of this research.
CONSENT FOR PUBLICATION
Not applicable.
AVAILABILITY OF DATA AND MATERIALS
The data supporting the findings of the article is available in the manuscript.
FUNDING
None.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
Declared none.

