All published articles of this journal are available on ScienceDirect.

High Titer of Antibody Against Pneumococcal IgA1 Protease in Healthy Individuals
Authors Info & Affiliations
Abstract
Background and Objectives:
Considering rising antibiotic resistance in various strains of Streptococcus pneumoniae, there is a need to find new immunogenic candidates for developing pneumococcal vaccines. Immunoglobulin A1 (IgA1) protease is one of the virulence factors playing an important role in the pathogenesis of S. pneumoniae infections. In the present study, we aimed to evaluate the titer of antibody against pneumococcal recombinant IgA1 protease in the serum of healthy humans.
Materials and Methods:
A part of the IgA1 protease gene (705 bp) from S. pneumonia ATCC 49619 was amplified by PCR and then digested using restriction enzymes and ligated by the pET28a expression vector. The recombinant protein was expressed in E. coli BL21 strain. Affinity chromatography was used to purify the protein. The titer of antibody against the recombinant protease was determined in healthy individuals in three age groups of <2, 2-40, and > 40 years using indirect Enzyme-Linked Immunosorbent Assay (ELISA).
Results:
The expression and purification of the IgA1 recombinant protease were successful. The concentration of the purified protein was determined as 1.013 mg/ml using the NanoDrop method. The titer of anti-recombinant IgA1 protease antibody (20, 40, 80 and 160) showed a significant correlation with age (p-value<0.05). According to our results, the antibody titer was desirable, especially in individuals over two years old.
Conclusion:
In the present study, desirable antibody titers against the pneumococcal recombinant IgA1 protease were seen in the three groups’ serum of healthy individuals. However, a significant correlation was not totally observed among groups.
1. INTRODUCTION
Streptococcus pneumoniae is a Gram-positive and extracellular invasive bacterium colonizing mucosal surfaces of upper airways in carriers and patients [1, 2]. The bacterium is an important cause of global morbidity and mortality, inflicting more deaths than any other bacterial infection. It causes mild respiratory tract infections such as otitis media and sinusitis to more severe diseases such as pneumonia, septicemia and meningitis [3, 4]. The main virulence factors involved in the pathogenesis of pneumococcal infections include Capsular Polysaccharide (CPS), Pneumolysin (Ply), Pneumococcal surface protein A (PspA), Choline-binding protein A (CbpA), CbpE, autolysin LytA, Immunoglobulin A1 (IgA1) protease, ZmpB, ZmpC, lipopeptides, lipoteichoic acid, and peptidoglycan fragments [5].
Over the past decades, the resistance of S. pneumoniae to various antimicrobial agents such penicillins has increased the successful hurdling treatment of the infection [6]. The best approach to reduce the mortality rate and treatment costs of S. pneumoniae infections seems to be designing an appropriate vaccine to prevent the disease. The bacterial polysaccharide capsule, which has been used to develop pneumococcal vaccines, is one of the most important antigenic determinants of this bacterium. There are currently two types of vaccines available for the disease: polysaccharide and conjugate vaccines. The polysaccharide vaccine, known as Pneumovax 23, consists of purified capsular antigens derived from 23 bacterial serotypes. The recent vaccine, however, is not effective in younger children. Unlike the Pneumovax 23, conjugate vaccines such as Prevnar13 and Synflorix promote high immunity in all age groups [7, 8].
Nowadays, in addition to encapsulated bacteria, non-encapsulated strains of S. pneumoniae are frequently isolated from many patients with pneumococcal infections. Therefore, current vaccines, which are generally based on capsular antigens, are not effective against the newly emerged infections [9]. For this reason, researchers have been investigating other proteins of the bacterium to develop new vaccines with the ability to induce high protection against all pneumococcal serotypes.
The IgA1 protease, as one of the most important pneumococcal proteins, facilitates the entry and colonization of the bacterium by destroying the mucosal IgA antibody [10]. The IgA1 protease of S. pneumoniae, as a metalloproteinase with a molecular weight of 215 KDa, breaks down the IgA1 at the hinge region (between Pro-227 and Thr-228) within the heavy chain of the antibody. IgA1 constitutes more than 90% of the IgA in the human airways. The cleavage of antigen-bound IgA1 creates fab fragments on the bacterial surface that inhibits phagocytosis, and consequently, the bacterial cells are not effectively neutralized [11]. Indeed, the decrease in pneumococcal colonization following the use of the capsular conjugate vaccine may be due to its induction of high IgG levels that are not targeted by the IgA1 protease [12, 13]. However, the main antibody-mediated defense in airways is related to the IgA class, and its opsonization role has been proven [14]. IgA1 protease is a conserved protein in S. pneumoniae serotypes and plays a main role in the control of the host immune response, it may be useful to apply in pneumococcal vaccines [15]. In the present study, the pneumococcal truncated IgA1 protease gene was cloned, expressed, and purified and then assessed the titer of its corresponding antibody in the serum of healthy volunteers.
2. METHODS
2.1. DNA Extraction, PCR and Cloning of Recombinant Protease Gene
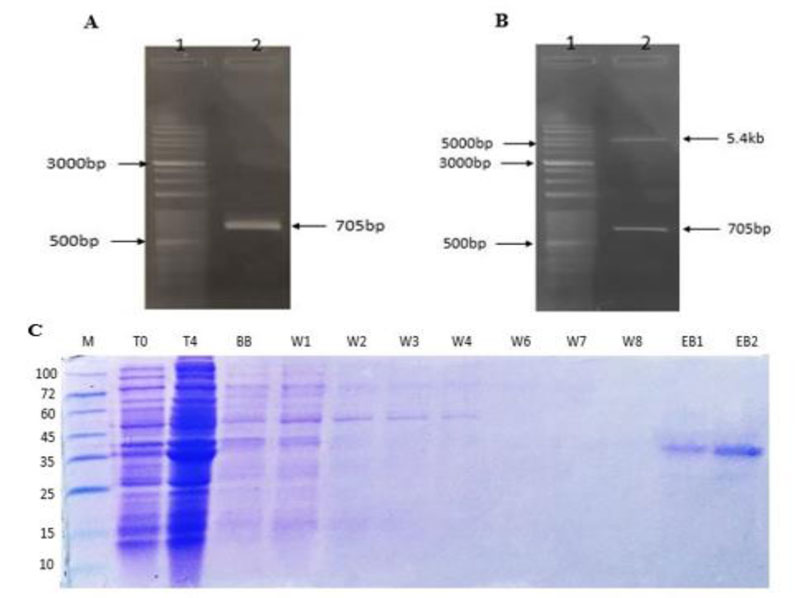
Genomic DNA was extracted from S. pneumoniae ATCC 49619 strain using a commercial DNA extraction Kit (FAVORGEN) according to the manufacturer’s instructions. Polymerase chain reaction (PCR) was carried out using 0.5 μM of forward primer (CCCA CAT ATG AAC GGA ACT GCA ACA AAA C), 0.5 μM of reverse primer (CCCA CTC GAG ACC ATC AAC ATG AAC TTG TT), 25 ng template DNA and 12.5μl of PCR-mastermix (2X). PCR product (705 bp) was purified using a DNA Gel Recovery Kit (PCR purification kit, GTP, Iran) and then double-digested by Nde-1 and Xho-1 restriction enzymes (Thermo Fisher Scientific, U.S) by a mixture containing 2 μl buffer O (10X), 3 μl Nde-1, 3μl Xho-1 and 1μg purified amplicon. The digested amplicons were ligated to the (predigested with Nde-1 and Xho-1 restriction enzymes) PET28a vector using T4 ligase enzyme (Takara, Japan). The ligation reaction was carried out for 16 h at 4°C, and the ligated product was transformed into E. coli DH5-α strain using the chemical CaCL2 transformation method [16]. The transformation suspension was cultured in LB agar containing kanamycin (1μg/ml) and incubated at 37°C for 24 h. Plasmid extraction was performed using a plasmid extraction kit (Bio Basic, Canada) according to the manufacturer’s instructions.
2.2. Recombinant Protein Expression, Purification and Confirmation
Recombinant PET28a (705 bp) protein was transformed into E. coli BL21 based on the chemical CaCl2 method, as described in the section above. The transformed bacterial cells were cultured in LB agar (Merck, Germany) containing kanamycin (1μg/ml) and incubated at 37°C for 24 h. The next day, a single colony was cultured in LB broth containing kanamycin (1μg/ml) and incubated at 37°C for 24 h. A 200 µl volume of the bacteria grown overnight was sub-cultured in LB broth supplemented with kanamycin (1μg/ml) and incubated at 37°C to reach the desired optical density (OD: 0.6 at 600 nm). In order to induce the expression of the recombinant protein, 1 mM Isopropyl β-d-1-thiogalactopyranoside (IPTG) was added to the culture of the transformed bacteria, and after 4 hours, the bacterial cell membranes were destroyed by sonication. The recombinant protein was purified by Ni-NTA affinity chromatography, and its concentration determined by reading ODs at 280 nm and 260 nm using a Nanodrop device and the following formula:
Protein concentration (mg/ml): (1.55 x OD280) - (0.76 x OD260)
The quality of the purified IgA protease recombinant protein was assessed by 10% SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE). Briefly, five μg of IgA protease recombinant protein was loaded into well of polyacrylamide gel and electrophoresed at 150V for 3 h. The gel was stained with coomassie brilliant blue for 30 min and destained with the destaining solution (100 ml deionized water, 100 ml methanol and 100 ml acetic acid). To confirm IgA protease recombinant protein, it was electrophoresed in 10% polyacrylamide gel and then transferred into polyvinylidene difluoride (PVDF) membrane. The membrane was blocked in blocking solution (Phosphate-Buffered Saline (PBS) containing 0.05% Tween 20 and 5% skimmed milk) at 4°C overnight and after washing with wash buffer (PBS containing 0.05% Tween 20), the membrane incubated with monoclonal anti-His tagged antibody conjugated to Horseradish Peroxidase (HRP) for 1 h. The membrane was washed 3 times with wash buffer and incubated with 3, 3’-Diaminobenzidine(DAB) substrate for 3 min.
2.3. Detection of Antibodies Against IgA1 Protease Recombinant Protein
Serum samples were obtained from sixty healthy people aged from <2, 2 to 40 and more than 40 years old. Antibodies against truncated-IgA1 protease were detected by indirect ELISA assay. One hundred μl of truncated-IgA1 protease recombinant protein (10 μg/mL) was added into 96-well polystyrene microplates (Cytomatin, Isfahan) and incubated at 4°C for 12 h. The wells were washed 3 times with washing buffer and blocked with 5% skim milk at 25°C for 1 h. The wells were washed 3 times with washing buffer, 100 μl from each serum at dilution of 1:10 to 1:160 added into the wells, and incubated at 25°C for 1h. The wells were washed 3 times with washing buffer, and 100 μl anti-human IgG conjugated with peroxidase (Cytomatin, Isfahan, Iran) was added into the wells and incubated at 25°C for 30 min. The wells were again washed 3 times with washing buffer and 100 μl 3,3′,5,5′-Tetramethylbenzidine (TMB) (Cytomatin Gene Co, Isfahan, Iran) substrate added into the wells. After 15 min, the reaction was terminated by 100 μl of 1N H2SO4 and optical density (OD) of samples was measured at 450-630 nm.
2.4. Statistical Analysis
The data was analyzed by SPSS version 16.0 software (SPSS, Inc., Chicago, IL). The participants were categorized into three age groups of <2, 2-40, and >40 years. One-way ANOVA (Tukey) test was used to compare the antibody titer among the three groups. A p-value less than 0.05 was regarded as statistically significant.
3. RESULTS
3.1. IgA1 Protease Gene Cloning, Purification and Confirmation of the Recombinant Protein
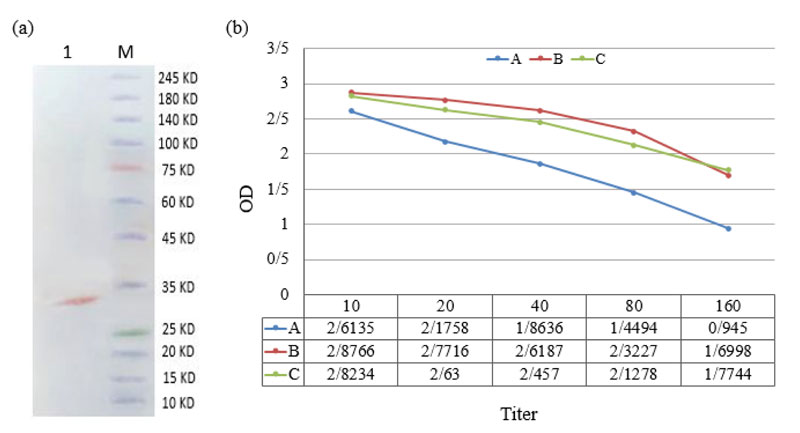
The expected PCR-product band was observed in agarose gel electrophoresis, as shown in Fig. (1A) . The cloning of the IgA1 protease gene was confirmed by double-digestion of the recombinant vector by Nde I and Xho I restriction enzymes ( Fig. 1B ), and the recombinant vector was optimally expressed in 1 mM concentration of IPTG after 4h induction. Purification of protein was successfully yielded 1.013 mg/ml truncated IgA protease with high purity ( Fig. 1C ). The purified IgA protease was confirmed by western blot analysis (Fig. 2a ).
3.2. Serological Analysis
Comparison of anti-recombinant IgA1 protease antibody titers among three groups showed a significant response of anti-recombinant IgA1 protease antibody between individuals >2 years and those who have <2 years old (P < 0.05). A significant highest antibody titers were observed in the group with 2-40-years old while the lowest titers belonged to children <2 years old (P < 0.05). One-way ANOVA (Tukey) test revealed a significant difference in the averages of antibody titers (20, 40, 80 and 160) between the groups of <2 and > 2 years old(P < 0.05) (Fig. 2b ).
4. DISCUSSION
IgA1 protease, which cleaves human IgA1, is one of the most important virulence factors produced by S. pneumoniae and several other invasive mucosal pathogens (e.g., Neisseria species and Haemophilus influenzae) [17]. Given the importance of streptococcal infections and the lack of an effective vaccine against all strains of S. pneumoniae [9], we here cloned, expressed, and purified a part of the pneumococcal IgA1 protease to assess its immunogenicity. For this purpose, antibody titer against the truncated recombinant IgA1 protease was determined in the serum of healthy individuals from three age groups. Our findings showed an appropriate level of the anti-recombinant IgA1protease antibody in healthy individuals, especially those aged 2-40 years old.
In a study by Romanello et al., the researchers cloned, expressed, and purified three segments of the S. pneumoniae IgA1 protease to investigate their function and immuno genicity. The results of the recent study showed a replacement of alanine with glutamate in the HEXXH motif (1605-1609) within the conserved domain of IgA protease. Considering the identification of antibodies against these segments of the IgA protease, it would be possible to develop vaccines against S. pneumoniae using inactive mutant IgA protease peptides [18]. In another study by Janoff et al., it was shown that the IgA protease could prevent removing of S. pneumoniae bacteria by human IgA in vivo [19]. The recent study supports the applicability of IgA protease as a potential candidate for designing vaccines against S. pneumoniae.
In the present study, the IgA1 proteolytic domain (corresponding to the 1465 to 2616 nucleoids of the corresponding gene) was cloned, expressed, and purified as a recombinant truncated protein. The serological analysis revealed a satisfactory antibody titer against this protease in healthy individuals, especially in those aged 2 to 40 years old. Probably, the suitable antibody titers against IgA protease in middle-age groups resulted from exposure to other bacterial colonization in the oral cavity that produces IgA protease and stimulates mucosal immunity. A study by Poulsen et al. showed that there is a 75 to 79% sequence similarity of IgA1 proteases between S. pneumoniae and S. oralis and S. mitis [20].
There was a significant association between the titer of anti-pneumococcal IgA1 protease antibody and age. In this regard, statistical analysis showed significant differences in antibody titers (20, 40, 80 and 160) between the two age groups of < 2 and 2-40years. The possible explanation for the lower antibody titers in the age group of < 2 years may be incomplete colonization of S. pneumoniae in young children and lack of maternal antibodies against IgA protease. Likewise, a reduction in the antibody titers in people over 40 years old can be related to the weakening of the immune system with age.
CONCLUSION
In the present study, desirable antibody titers against the pneumococcal recombinant IgA1 protease were seen in the three groups’ serum of healthy individuals. However, a significant correlation was not totally observed among groups. A high titer of antibody against pneumococcal recombinant IgA1 protease in healthy groups may be due to sustained stimulation of the immune system by IgA1 proteases produced by bacteria naturally found in the oral cavity.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
Not applicable.
HUMAN AND ANIMAL RIGHTS
No animal or human were the basis of this research.
CONSENT FOR PUBLICATION
Not applicable.
AVAILABILITY OF DATA AND MATERIALS
Not applicable.
FUNDING
None.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
Declared none.

